Дофами́н (допами́н[1], DA) — нейромедиатор, вырабатываемый в мозгу людей и животных. Также гормон, вырабатываемый мозговым веществом надпочечников и другими тканями (например, почками), но в подкорку мозга из крови этот гормон почти не проникает. По химической структуре дофамин относят к катехоламинам. Дофамин является биохимическим предшественником норадреналина (и адреналина).
Нейромедиатор
Дофамин является одним из химических факторов внутреннего подкрепления (ФВП) и служит важной частью «системы поощрения» мозга, поскольку вызывает чувство удовольствия (или удовлетворения) чем влияет на процессы мотивации и обучения[2][3]. Дофамин естественным образом вырабатывается в больших количествах во время позитивного, по субъективному представлению человека, опыта — к примеру, секса, приёма вкусной пищи, приятных телесных ощущений, а также стимуляторов, ассоциированных с ними
[4][5]. Нейробиологические эксперименты показали, что даже воспоминания о позитивном поощрении могут увеличить уровень дофамина[6][2][3][7], поэтому данный нейромедиатор используется мозгом для оценки и мотивации, закрепляя важные для выживания и продолжения рода действия[8].
Дофамин играет немаловажную роль в обеспечении когнитивной деятельности. Активация дофаминергической передачи необходима при процессах переключения внимания человека с одного этапа когнитивной деятельности на другой. Таким образом, недостаточность дофаминергической передачи приводит к повышенной инертности больного, которая клинически проявляется замедленностью когнитивных процессов (брадифрения) и персеверациями. Данные нарушения являются наиболее типичными когнитивными симптомами болезней с дофаминергической недостаточностью — например, болезни Паркинсона [9].
Как и у большинства нейромедиаторов, у дофамина существуют синтетические аналоги, а также стимуляторы его выделения в мозге. В частности, многие наркотики увеличивают выработку и высвобождение дофамина в мозге в 5—10 раз, что позволяет людям, которые их употребляют, получать чувство удовольствия искусственным образом[10][11][12]. Так, амфетамин напрямую стимулирует выброс дофамина, воздействуя на механизм его транспортировки.[13] Другие наркотики, например, кокаин и психостимуляторы, блокируют естественные механизмы обратного захвата дофамина, увеличивая его концентрацию в синаптическом пространстве[14]. Морфий и никотин имитируют действие натуральных нейромедиаторов[14], а алкоголь блокирует действие антагонистов дофамина[15]. Если пациент продолжает перестимулировать свою «систему поощрения», постепенно мозг адаптируется к искусственно повышаемому уровню дофамина, производя меньше гормона и снижая количество рецепторов в «системе поощрения»
[16], что побуждает наркомана увеличивать дозу для получения прежнего эффекта. Дальнейшее развитие химической толерантности может постепенно привести к очень тяжёлым изменениям в нейронах и других структурах мозга, а в долговременной перспективе потенциально нанести серьёзный ущерб здоровью мозга[17].
Для лечения болезни Паркинсона часто используют агонисты дофаминовых рецепторов (прамипексол, бромокриптин, перголид и др.): на сегодняшний день это самая многочисленная группа противопаркинсонических средств[18]. Некоторые из антидепрессантов также обладают дофаминергической активностью[19].
Существуют и лекарственные препараты, блокирующие дофаминергическую передачу, например такие антипсихотические средства, как аминазин, галоперидол, рисперидон, клозапин и др. Резерпин блокирует накачку дофамина в пресинаптические везикулы.
При таких психических заболеваниях, как шизофрения и обсессивно-компульсивное расстройство, отмечается повышенная дофаминергическая активность в некоторых структурах мозга (при шизофрении отмечается вдобавок пониженная активность дофамина в мезокортикальном дофаминовом пути [20][21][22][23] и префронтальной коре[20][24][23]), а паркинсонизм связан с пониженным содержанием дофамина в нигростриарном пути[24][25][26][27]. Со снижением уровня дофамина в подкорковых образованиях и передних отделах головного мозга связывают также процесс нормального старения[28].
Биосинтез
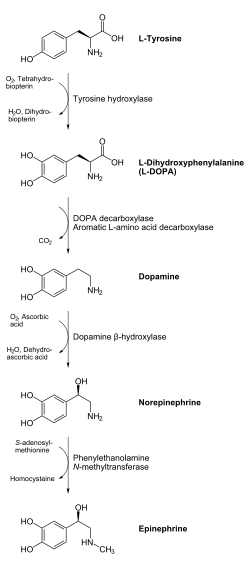
Предшественником дофамина является L-тирозин (он синтезируется из фенилаланина), который гидроксилируется ферментом тирозингидроксилазой с образованием L-ДОФА, которая, в свою очередь, декарбоксилируется с помощью фермента L-ДОФА-декарбоксилазы и превращается в дофамин. Этот процесс происходит в цитоплазме нейрона.
Рецепторы
Постсинаптические дофаминовые рецепторы относятся к семейству GPCR. Существует по меньшей мере пять различных подтипов дофаминовых рецепторов — D1—5. Рецепторы D1 и D5 обладают довольно значительной гомологией и сопряжены с белком GS, который стимулирует аденилатциклазу, вследствие чего их обычно рассматривают совместно как D-1-подобные рецепторы. Остальные рецепторы подсемейства подобны D2 и сопряжены с Gi-белком, который ингибирует аденилатциклазу, вследствие чего их объединяют под общим названием D-2-подобные рецепторы. Таким образом, дофаминовые рецепторы играют роль модуляторов долговременной потенциации[29].
Участие во «внутреннем подкреплении» принимают D2 и D4 рецепторы.
В больших концентрациях дофамин также стимулирует α- и β-адренорецепторы. Влияние на адренорецепторы связано не столько с прямой стимуляцией адренорецепторов, сколько со способностью дофамина высвобождать норадреналин из гранулярных пресинаптических депо, то есть оказывать непрямое адреномиметическое действие.
«Круговорот» дофамина
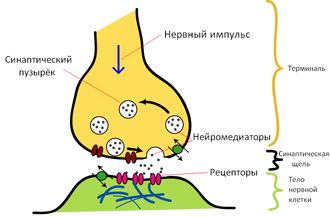
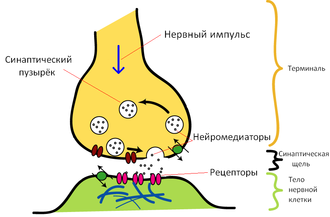
Основные элементы синапса
Синтезированный нейроном дофамин накапливается в дофаминовых везикулах (т. н. «синаптическом пузырьке»). Этот процесс является протон-сопряжённым транспортом. В везикулу с помощью протон-зависимой АТФазы закачиваются ионы H+. При выходе протонов по градиенту в везикулу поступают молекулы дофамина.
Далее дофамин выводится в синаптическую щель. Часть его участвует в передаче нервного импульса, воздействуя на клеточные D-рецепторы постсинаптической мембраны, а часть возвращается в пресинаптический нейрон с помощью обратного захвата. Ауторегуляция выхода дофамина обеспечивается D2 и D3 рецепторами на мембране пресинаптического нейрона. Обратный захват производится транспортером дофамина. Вернувшийся в клетку медиатор расщепляется с помощью моноаминооксидазы (МАО) и, далее, альдегиддегидрогеназы и катехол-О-метил-трансферазы до гомованилиновой кислоты.
Участие в системе поощрения
Лабораторная крыса в специальном ящике нажимает рычаг. К голове животного прикреплены стимуляторы.
В фундаментальном исследовании 1954 года канадские учёные Джеймс Олдс и его коллега Питер Милнер обнаружили, что если имплантировать электроды в определённые участки мозга, особенно в средний узел переднего мозга, то крысу можно приучить нажимать рычаг в клетке, включающий стимуляцию низковольтными разрядами электричества[30]. Когда крысы научились стимулировать этот участок, они нажимали рычаг до тысячи раз в час[30]. Это дало основание предположить, что стимулируется центр наслаждения. Один из главных путей передачи нервных импульсов в этом участке мозга — дофаминовый, поэтому исследователи выдвинули версию, что главное химическое вещество, связанное с удовольствием, — это дофамин. В дальнейшем это предположение было подтверждено радионуклидными томографическими сканерами и открытием антипсихотиков (лекарственных средств, подавляющих продуктивные симптомы шизофрении) [31].
Дофаминергическая система
Из всех нейронов ЦНС только около семи тысяч вырабатывают дофамин. Известно несколько дофаминовых ядер, расположенных в мозге. Это дугообразное ядро (лат. nucleus arcuatum), дающее свои отростки в срединное возвышение гипоталамуса. Дофаминовые нейроны чёрной субстанции посылают аксоны в стриатум (хвостатое и чечевицеобразное ядро). Нейроны, находящиеся в области вентральной покрышки, дают проекции к лимбическим структурам и коре.
Основные дофаминовые пути.
Основными дофаминовыми путями являются:
Тела нейронов нигростриатного, мезокортикального и мезолимбического трактов образуют комплекс нейронов чёрной субстанции (англ.)русск. и вентрального поля покрышки. Аксоны этих нейронов идут вначале в составе одного крупного тракта (медиального пучка переднего мозга), а далее расходятся в различные мозговые структуры. Некоторые авторы объединяют мезокортикальную и мезолимбическую подсистемы в единую систему, однако более обоснованно выделение мезокортикальной и мезолимбической подсистем соответственно проекциям в лобную кору и лимбические структуры мозга
[32].
В экстрапирамидной системе дофамин играет роль стимулирующего нейромедиатора, способствующего повышению двигательной активности, уменьшению двигательной заторможенности и скованности, снижению гипертонуса мышц. Физиологическими антагонистами дофамина в экстрапирамидной системе являются ацетилхолин и ГАМК.
Другие подсистемы
Выделяют также тубероинфундибулярный путь (лимбическая система — гипоталамус — гипофиз), инцертогипоталамический, диенцефалоспинальный и ретинальный[32] (иногда, вдобавок к этому, перивентрикулярную
и ольфакторную системы[33]). Данная дифференциация не является абсолютной, поскольку проекции дофаминергических нейронов разных трактов «перекрываются»; кроме того, в мозге отмечается и диффузное распределение дофаминергических элементов (отдельных клеток с отростками)[32].
В гипоталамусе и гипофизе дофамин играет роль естественного тормозного нейромедиатора, угнетающего секрецию ряда гормонов. При этом угнетающее действие на секрецию разных гормонов реализуется при разных концентрациях дофамина, что обеспечивает высокую специфичность регуляции. Наиболее чувствительна к тормозящему действию дофаминергических сигналов секреция пролактина, в меньшей степени — секреция соматолиберина и соматотропина, в ещё меньшей — секреция кортиколиберина и кортикотропина и в совсем малой степени — секреция тиролиберина и тиротропина. Секреция гонадотропинов и гонадолиберина не угнетается дофаминергическими сигналами.
Ввиду чувствительности некоторых гормональных подсистем к уровню дофамина препараты-дофаминомиметики, усиливающие его синтез, могут применяться в качестве терапии при гормональных заболеваниях. Например, дофаминомиметики назначают при гиперпролактинемии и при болезни Паркинсона.
Дофамин и другие нейромедиаторы
Дофаминергические подсистемы находятся под контролем или сами контролируют норадренергические, серотонинергические, ГАМК-ергические, холинергические, мелатонинергические, глутаматергические, пептидергические системы. ГАМК-ергические и серотонинергические системы находятся в антагонистических отношениях с дофаминергической системой, а норадренергическая и дофаминергическая системы в различных функциональных состояниях функционируют однонаправленно: как в период бодрствования, так и в период сна. Взаимодействия дофаминергической и холинергической системы сложны, в условиях патологических процессов активность этих систем неоднозначна[33].
Гормон
Дофамин обладает рядом физиологических свойств, характерных для адренергических веществ.
Дофамин вызывает повышение сопротивления периферических сосудов (менее сильное, чем под влиянием норадреналина). Он повышает систолическое артериальное давление в результате стимуляции α-адренорецепторов. Также дофамин увеличивает силу сердечных сокращений в результате стимуляции β-адренорецепторов. Увеличивается сердечный выброс. Частота сердечных сокращений увеличивается, но не так сильно, как под влиянием адреналина.
Потребность миокарда в кислороде под влиянием дофамина повышается, однако в результате увеличения коронарного кровотока обеспечивается повышенная доставка кислорода.
В результате специфического связывания с дофаминовыми рецепторами почек дофамин уменьшает сопротивление почечных сосудов, увеличивает в них кровоток и почечную фильтрацию. Наряду с этим повышается натрийурез. Происходит также расширение мезентериальных сосудов. Этим действием на почечные и мезентериальные сосуды дофамин отличается от других катехоламинов (норадреналина, адреналина и др.). Однако в больших концентрациях дофамин может вызывать сужение почечных сосудов.
Дофамин ингибирует также синтез альдостерона в коре надпочечников, понижает секрецию ренина почками, повышает секрецию простагландинов тканью почек.
Дофамин тормозит перистальтику желудка и кишечника, вызывает расслабление нижнего пищеводного сфинктера и усиливает желудочно-пищеводный и дуодено-желудочный рефлюкс. В ЦНС дофамин стимулирует хеморецепторы триггерной зоны и рвотного центра и тем самым принимает участие в осуществлении акта рвоты.
Через гематоэнцефалический барьер дофамин мало проникает, и повышение уровня дофамина в плазме крови оказывает малое влияние на функции ЦНС, за исключением действия на находящиеся вне гематоэнцефалического барьера участки, такие как триггерная зона.
Повышение уровня дофамина в плазме крови происходит при шоке, травмах, ожогах, кровопотерях, стрессовых состояниях, при различных болевых синдромах, тревоге, страхе, стрессе. Дофамин играет роль в адаптации организма к стрессовым ситуациям, травмам, кровопотерям и др.
Также уровень дофамина в крови повышается при ухудшении кровоснабжения почек или при повышенном содержании ионов натрия, а также ангиотензина или альдостерона в плазме крови. По-видимому, это происходит вследствие повышения синтеза дофамина из ДОФА в ткани почек при их ишемии или при воздействии ангиотензина и альдостерона. Вероятно, этот физиологический механизм служит для коррекции ишемии почек и для противодействия гиперальдостеронемии и гипернатриемии.
Патологии
Наиболее известными патологиями, связанными с дофамином, являются шизофрения и паркинсонизм, а также обсессивно-компульсивное расстройство. Различные независимые исследования показали, что многие лица, страдающие шизофренией, имеют повышенную дофаминергическую активность в некоторых структурах мозга[24][25][27] (и пониженную дофаминергическую активность в мезокортикальном пути[20][23][21][22] и префронтальной коре[20][23][24]). Для лечения шизофрении применяются антипсихотики (нейролептики), которые блокируют рецепторы дофамина (преимущественно D2-типа) и варьируются в степени аффинности к другим значимым нейромедиаторным рецепторам[34]. Типичные антипсихотики в основном подавляют рецепторы D2, а новые атипичные антипсихотики и некоторые из типичных воздействуют одновременно на целый ряд нейромедиаторных рецепторов: дофамина, серотонина, гистамина, ацетилхолина и других[34].
Предполагается, что снижение уровня дофамина в мезокортикальном пути связано с негативными симптомами шизофрении[20] (сглаживание аффекта, апатия, бедность речи, ангедония, уход из общества[23]), а также с когнитивными расстройствами[20] (дефициты внимания, рабочей памяти, исполнительных функций[23]). Антипсихотическое действие нейролептиков, то есть их способность редуцировать продуктивные нарушения — бред, галлюцинации, психомоторное возбуждение — связывают с угнетением дофаминергической передачи в мезолимбическом пути[22]. Нейролептики также угнетают дофаминергическую передачу и в мезокортикальном пути[35], что при длительной терапии может приводить к усилению негативных нарушений[36].
Паркинсонизм связан с пониженным содержанием дофамина в нигростриарном пути. Наблюдается при разрушении чёрной субстанции, патологии D-1-подобных рецепторов. С угнетением дофаминергической передачи в нигростриарной системе связывают и развитие экстрапирамидных побочных эффектов при приёме антипсихотиков[22]: лекарственного паркинсонизма, дистонии, акатизии, поздней дискинезии и др.
С нарушением дофаминергической системы связывают такие расстройства, как ангедония, депрессия, деменция, патологическая агрессивность, фиксация патологических влечений, синдром персистирующей лактореи-аменореи, импотенция, акромегалия, синдром беспокойных ног и периодических движений в конечностях[33].
Процесс старения
По данным исследований, процесс старения проявляется уменьшением объема и массы головного мозга и уменьшением числа синаптических связей; кроме уменьшения числа церебральных рецепторов, имеет место и медиаторная церебральная недостаточность. С возрастом уменьшается количество и плотность дофаминовых D2-рецепторов стриатума, снижается концентрация дофамина в подкорковых образованиях головного мозга. Клиническими проявлениями этих изменений являются обеднение мимики, некоторая общая замедленность, сгорбленная, старческая поза, укорочение длины шага.
«Дофамин-чувствительные» изменения отмечаются также в когнитивной сфере: с возрастом снижается быстрота реакции, становится труднее усваивать и реализовывать новую программу действия, снижается уровень внимания, объём оперативной памяти. При отсутствии органической патологии возрастные когнитивные изменения не приводят к дезадаптации пожилых людей и позволяют поддерживать привычный ритм социальной активности[28].
См. также
Примечания
↑ В русской терминологии принято название «дофамин». Вариант «допамин» представляет собой прямую транслитерацию принятого в западной литературе термина dopamine и отсутствует в русских словарях. Различие объясняется разницей в сокращениях, возникшей из-за соответствия ph = ф: dihydroxyphenylalanine = DOPA, дигидроксифенилаланин = ДОФА.
↑ 12 It’s all about dopamine. Архивировано из первоисточника 22 августа 2011.
↑ 12 Biology of Happiness. Архивировано из первоисточника 22 августа 2011.
↑ Допамин — удовольствие от жизни. Архивировано из первоисточника 3 февраля 2012.
↑ Ваш мозг во время секса. Архивировано из первоисточника 22 августа 2011.
↑ Удовольствие: дофамин. Архивировано из первоисточника 24 января 2012.
↑ Кокаиновые наркоманы могут получать удовольствие только от одного ожидания этого удовольствия. Архивировано из первоисточника 24 января 2012.
↑ Dopamine and desire. Архивировано из первоисточника 22 августа 2011.
↑Захаров В.В., Яхно Н.Н. Когнитивные расстройства в пожилом и старческом возрасте: Методическое пособие для врачей. — Москва, 2005.
↑ Мозг и наркотики. Архивировано из первоисточника 22 августа 2011.
↑ Your Brain on Drugs: Dopamine and Addiction. Архивировано из первоисточника 22 августа 2011.
↑ Dopamine — A Sample Neurotransmitter. Архивировано из первоисточника 22 августа 2011.
↑ A Mechanism for Amphetamine-Induced Dopamine Overload.
↑ 12 HOW DRUGS AFFECT NEUROTRANSMITTERS. Архивировано из первоисточника 22 августа 2011.
↑ Мужчины чаще становятся алкоголиками из-за дофамина, выяснили ученые. Архивировано из первоисточника 22 августа 2011.
↑ Addiction and Dopamine (D2) Receptor Levels (2006). Архивировано из первоисточника 22 августа 2011.
↑ The Science Behind Drug Use and Addiction. Архивировано из первоисточника 22 августа 2011.
↑Катунина Е. А. Агонисты дофаминовых рецепторов и проблема эквивалентности доз // Неврология. — 2010. — № 2.
↑Shelton RC, Tomarken AJ Can Recovery From Depression Be Achieved? // American Psychiatric Association Psychiatr Serv. — November 2001. — № 52. — С. 1469—1478. Перевод: Можно ли достичь выздоровления при депрессии? // Обзор современной психиатрии. — November 2004. — В. 21.
↑ 123456 Abi-Dargham A, Moore H (October 2003). «Prefrontal DA transmission at D1 receptors and the pathology of schizophrenia». Neuroscientist9 (5): 404–16. DOI:10.1177/1073858403252674. PMID 14580124.
↑ 12 Lieberman JA (2004). «Dopamine partial agonists: a new class of antipsychotic». CNS Drugs18 (4): 251–67. PMID 15015905.
↑ 1234 Справочное руководство по психофармакологическим и противоэпилептическим препаратам, разрешенным к применению в России / Под ред. С. Н. Мосолова. — 2-е, перераб. — М.: «Издательство БИНОМ», 2004. — С. 17. — 304 с. — 7000 экз. — ISBN 5-9518-0093-5
↑ 123456Abi-Dargham A The Dopamine Hypothesis of Schizophrenia. Schizophrenia Research Forum (5 December 2005). Архивировано из первоисточника 3 февраля 2012. Проверено 26 сентября 2011. Перевод: Дофаминовая гипотеза шизофрении (15 August 2008). Архивировано из первоисточника 3 февраля 2012.
↑ 1234 The Dopamine Hypothesis of Schizophrenia: Version III—The Final Common Pathway. Архивировано из первоисточника 23 августа 2011.
↑ 12 Increased Striatal Dopamine Transmission in Schizophrenia: Confirmation in a Second Cohort. Архивировано из первоисточника 23 августа 2011.
↑ Presynaptic Regulation of Dopamine Transmission in Schizophrenia. Архивировано из первоисточника 23 августа 2011.
↑ 12 Dysconnection in Schizophrenia: From Abnormal Synaptic Plasticity to Failures of Self-monitoring. Архивировано из первоисточника 23 августа 2011.
↑ Mehta M.A., Riedel W.J. Dopaminergic Enhancement of Cognitive Function. // Curr. Pharm. Des. 2006. v. 12. pp. 2487—2500.
↑ 12 Британская энциклопедия, см. раздел «Reward and punishment». Архивировано из первоисточника 3 февраля 2012.
↑ The Functional Neuroanatomy of Pleasure and Happiness. Архивировано из первоисточника 3 февраля 2012.
↑ 123Орловская Д. Д. Нейрохимические системы мозга // Общая психиатрия / Под ред. А. С. Тиганова. — Москва, 2006.
↑ 123Левин Я.И. Нейрохимическая медицина. Часть 1. Церебральные дофаминергические системы // Современная терапия психических расстройств. — 2008. — № 1.
↑ 12 A Roadmap to Key Pharmacologic Principles in Using Antipsychotics. PubMed (USA).
↑ Basu D, Marudkar M, Khurana H (2000). «Abuse of neuroleptic drug by psychiatric patients» 54 (2): 59—62. PMID 11271726.
↑Бородин В. И. Сперидан (рисперидон) при терапии шизофренических психозов // Психиатрия и психофармакотерапия. — 2006. — В. 8 (4).
Литература
Ашмарин И. П., Ещенко Н. Д., Каразеева Е. П. Нейрохимия в таблицах и схемах. — М.: Экзамен, 2007.
Васильев В. Н. Диагностика и терапия инкурабельных нервных и психических заболеваний допаминовой этиологии. Биокоррекция Васильева. — М.: Медиакит, 2009. — 247 с. — ISBN 978-5-9901746-1-0
Почки:JGA (ренин) · перитубулярные клетки (EPO) · кальцитриол · простагландин
Сердце: натрийуретический пептид (ANP, BNP)
Шаблон:АТХ код C01
Дофаминовый рецептор — Википедия
Дофаминовые рецепторы — класс трансмембранных метаботропных G-белок-связанных клеточных рецепторов, играющих важную роль в функционировании центральной нервной системы позвоночных. Основной эндогенный лиганд-агонист этих рецепторов — дофамин. Дофаминовые рецепторы участвуют в процессах мотивации, обучения, тонкой моторной координации, модулирования нейроэндокринных сигналов. Этот класс включает пять типов рецепторов: D1, D2, D3, D4 и D5[1].
Изменение дофаминергической функции отмечается в ряде нейрологических и психических расстройств, а сами рецепторы являются мишенями для множества лекарственных препаратов. Подавляющее большинство антипсихотиков — антагонисты рецепторов дофамина, а психостимуляторы зачастую косвенно их активируют.
Известно пять генов человека, кодирующих рецепторы дофамина. По структурным, биохимическим и фармакологическим характеристикам соответствующие рецепторы подразделяют на D1-подобные (D1, D5) и D2-подобные (D2, D3, D4). Впервые эти два класса рецепторов были выделены в 1979 на основании того, что только D1-подобные рецепторы активируют аденилатциклазу[2]. Рецепторы группы D2 её, напротив, ингибируют[3].
Предполагается наличие рецепторов D6 и D7, но их существование пока не доказано.
Альтернативная классификация, предложенная в 1983[4],
подразделяет рецепторы по их эффектам: активация рецепторов группы DA1 вызывает релаксацию мышц и расширение сосудов; для этих рецепторов (R)-сульпирид является сильным антагонистом, апоморфин — слабым агонистом, а домперидон на них не действует. Активация рецепторов DA2 ингибирует действие норадреналина, апоморфин — их сильный агонист, а сильные антагонисты — (S)-сульпирид и домперидон. Дофаминовые рецепторы центральной нервной системы, по-видимому, относятся к этому классу.[5]
D1-подобные рецепторы
Как было сказано выше, к D1-подобным рецепторам относятся рецепторы D1 и D5. Характерной особенностью рецепторов этого класса является то, что они активируют G-белки семейства Gαs/olf, которые в свою очередь активируют аденилатциклазу[6].
D1-подобные рецепторы обнаруживаются только на постсинаптических мембранах клеток, чувствительных к дофамину. Гены рецепторов этого класса не содержат интронов, поэтому D1- и D5-рецепторы существуют в единственном сплайс-варианте[7][8].
D2-подобные рецепторы[править | править код]
К D2-подобным рецепторам относятся рецепторы D2, D3 и D4. Эти рецепторы связываются с G-белками семейства Gαi/o и поэтому ингибируют аденилатциклазу[3][1]. В отличие от D1-подобных, рецепторы D2 и D3 присутствуют не только на постсинаптических мембранах клеток, чувствительных к дофамину, но и на пресинаптических мембранах дофаминергических нейронов. Гены D2-подобных рецепторов содержат интроны: в гене рецептора D2 обнаружено 7 интронов, в гене D3 — 5, а в гене D4 — 3 (гены человека)[9][10][11]. Известно, что рецепторы D2 и D3 существуют в нескольких формах, что является результатом альтернативного сплайсинга их пре-мРНК[12][13][14]. Структурно D2-подобные рецепторы отличаются тем, что их С-концевые домены в 7 раз короче, чем у D1-подобных рецепторов[15].
Дофаминовые рецепторы присутствуют как в центральной нервной системе, так в периферических органах. Относительная доля дофаминергических нейронов в головном мозге невелика (менее 1/100 000 всех нейронов)[15]. Эти нейроны формируют несколько основных дофаминергических путей: нигростриарный, мезолимбический, мезокортикальный и тубероинфундибулярный[16].
Дофаминовый рецептор D1 является самым широко распространённым дофаминовым рецептором в головном мозге, он синтезируется в большем количестве чем другие рецепторы. Он обнаруживается в высокой концентрации в нигростриарном, мезолимбическом и мезокортикальном путях, а именно в лобных долях, полосатом теле, чёрной субстанции, прилежащем ядре, обонятельном бугорке и миндалевидном теле. Также в меньшей концентрации он присутствует в гиппокампе, мозжечке, таламической и гипоталамической областях[17][18][19][20].
Рецептор D2 в высокой концентрации присутствует в полосатом теле, обонятельном бугорке, прилежащем ядре, чёрной субстанции, гипоталамусе, вентральной области покрышки и миндалевидном теле, то есть примерно в тех же участках мозга, где обнаруживается и рецептор D1[21][22][16]. Тем не менее, дополнительные исследования помогли установить, что только 5—15 % проекционных нейронов дорсальной части полосатого тела экспрессируют оба рецептора одновременно. Остальные нейроны могут быть разделены на две группы, в зависимости от того, какой из рецепторов они содержат[1].
Рецептор D3 имеет более узкий профиль распространения, чем рецепторы, описанные выше. В наибольшей концентрации он присутствует в прилежащем ядре, обонятельном бугорке и островках Калеха. В существенно более низких концентрациях рецептор D3 обнаруживается в компактной части чёрной субстанции, вентральной области покрышки и мозжечке[23][24][25].
Уровень экспрессии рецептора D4 в мозге существенно ниже, чем рецептора D2. Доказано, что рецептор D4 присутствует в коре больших полушарий, гиппокампе, полосатом и миндалевидном телах[15].
Рецептор D5 синтезируется в небольшом количестве в разных участках мозга: в пирамидальных нейронах префронтальной коры, поясной коре, энторинальной коре, чёрной субстанции, зубчатой извилине, гиппокампе и гипоталамусе[1].
Все пять типов рецепторов дофамина встречаются и за пределами головного мозга. Так рецепторы D1, D2 и D4 были обнаружены в сетчатке, а рецептор D2 — в гипофизе[26][27][28]. Дофаминовые рецепторы синтезируются в разных пропорциях в клетках почек, надпочечников, симпатических ганглиев, кровеносных сосудов, сердца и пищеварительного тракта[1].
↑ 12345Beaulieu J. M., Gainetdinov R. R. The physiology, signaling, and pharmacology of dopamine receptors (англ.) // Pharmacol. Rev. (англ.)русск. : journal. — 2011. — Vol. 63, no. 1. — P. 182—217. — DOI:10.1124/pr.110.002642. — PMID 21303898.
↑ Kebabian J. W., Calne D. B. Multiple receptors for dopamine (англ.) // Nature. — 1979. — Vol. 277, no. 5692. — P. 93—96. — DOI:10.1038/277093a0. — PMID 215920.
↑ 12Borgundvaag B., George S. R. Dopamine inhibition of anterior pituitary adenylate cyclase is mediated through the high-affinity state of the D2 receptor (англ.) // Life Sci. (англ.)русск. : journal. — 1985. — Vol. 37, no. 4. — P. 379—386. — PMID 4040201.
↑ Goldberg L., Kohli J. Peripheral dopamine receptors: a classification based on potency series and specific antagonism (англ.) // Trends Pharm. Sci. : journal. — 1983. — Vol. 4. — P. 64.
↑ Альберт А. Избирательная токсичность = Selective toxicity / Под ред. проф. В. А. Филова. — М.: Медицина, 1989. — Т. 1. — С. 228. — 432 с. — ISBN 412-26010-7.
↑ Hervé D., Lévi-Strauss M., Marey-Semper I., Verney C., Tassin J. P., Glowinski J., Girault J. A. G(olf) and Gs in rat basal ganglia: possible involvement of G(olf) in the coupling of dopamine D1 receptor with adenylyl cyclase (англ.) // J. Neurosci. (англ.)русск. : journal. — 1993. — Vol. 13, no. 5. — P. 2237—2248. — PMID 8478697.
↑ Sunahara R. K., Niznik H. B., Weiner D. M., Stormann T. M., Brann M. R., Kennedy J. L., Gelernter J. E., Rozmahel R., Yang Y. L., Israel Y., et al. Human dopamine D1 receptor encoded by an intronless gene on chromosome 5 (англ.) // Nature : journal. — 1990. — Vol. 347, no. 6288. — P. 80—83. — DOI:10.1038/347080a0. — PMID 1975640.
↑ Grandy D. K., Zhang Y. A., Bouvier C., Zhou Q. Y., Johnson R. A., Allen L., Buck K., Bunzow J. R., Salon J., Civelli O. Multiple human D5 dopamine receptor genes: a functional receptor and two pseudogenes (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1991. — Vol. 88, no. 20. — P. 9175—9179. — PMID 1833775.
↑ Grandy D. K., Marchionni M. A., Makam H., Stofko R. E., Alfano M., Frothingham L., Fischer J. B., Burke-Howie K. J., Bunzow J. R., Server A. C., et al. Cloning of the cDNA and gene for a human D2 dopamine receptor (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1989. — Vol. 86, no. 24. — P. 9762—9766. — PMID 2532362.
↑ Sokoloff P., Giros B., Martres M. P., Bouthenet M. L., Schwartz J. C. Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics (англ.) // Nature : journal. — 1990. — Vol. 347, no. 6289. — P. 146—151. — DOI:10.1038/347146a0. — PMID 1975644.
↑ Van Tol H. H., Bunzow J. R., Guan H. C., Sunahara R. K., Seeman P., Niznik H. B., Civelli O. Cloning of the gene for a human dopamine D4 receptor with high affinity for the antipsychotic clozapine (англ.) // Nature : journal. — 1991. — Vol. 350, no. 6319. — P. 610—614. — DOI:10.1038/350610a0. — PMID 1840645.
↑ Giros B., Sokoloff P., Martres M. P., Riou J. F., Emorine L. J., Schwartz J. C. Alternative splicing directs the expression of two D2 dopamine receptor isoforms (англ.) // Nature : journal. — 1989. — Vol. 342, no. 6252. — P. 923—926. — DOI:10.1038/342923a0. — PMID 2531847.
↑ Monsma FJ Jr, McVittie L. D., Gerfen C. R., Mahan L. C., Sibley D. R. Multiple D2 dopamine receptors produced by alternative RNA splicing (англ.) // Nature : journal. — 1989. — Vol. 342, no. 6252. — P. 926—929. — DOI:10.1038/342926a0. — PMID 2480527.
↑ Giros B., Martres M. P., Pilon C., Sokoloff P., Schwartz J. C. Shorter variants of the D3 dopamine receptor produced through various patterns of alternative splicing (англ.) // Biochem. Biophys. Res. Commun. (англ.)русск. : journal. — 1991. — Vol. 176, no. 3. — P. 1584—1592. — DOI:10.1016/0006-291X(91)90469-N. — PMID 2039532.
↑ 123Rondou P., Haegeman G., Van Craenenbroeck K. The dopamine D4 receptor: biochemical and signalling properties (англ.) // Cell. Mol. Life Sci. : journal. — 2010. — Vol. 67, no. 12. — P. 1971—1986. — DOI:10.1007/s00018-010-0293-y. — PMID 20165900.
↑ 12Missale C., Nash S. R., Robinson S. W., Jaber M., Caron M. G. Dopamine receptors: from structure to function (англ.) // Physiol. Rev. (англ.)русск. : journal. — 1998. — Vol. 78, no. 1. — P. 189—225. — PMID 9457173.
↑ Fremeau RT Jr, Duncan G. E., Fornaretto M. G., Dearry A., Gingrich J. A., Breese G. R., Caron M. G. Localization of D1 dopamine receptor mRNA in brain supports a role in cognitive, affective, and neuroendocrine aspects of dopaminergic neurotransmission (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1991. — Vol. 88, no. 9. — P. 3772—3776. — PMID 2023928.
↑ Weiner D. M., Levey A. I., Sunahara R. K., Niznik H. B., O’Dowd B. F., Seeman P., Brann M. R. D1 and D2 dopamine receptor mRNA in rat brain (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1991. — Vol. 88, no. 5. — P. 1859—1863. — PMID 1825729.
↑ Hurd Y. L., Suzuki M., Sedvall G. C. D1 and D2 dopamine receptor mRNA expression in whole hemisphere sections of the human brain (англ.) // J. Chem. Neuroanat. : journal. — 2001. — Vol. 22, no. 1—2. — P. 127—137. — DOI:10.1016/S0891-0618(01)00122-3. — PMID 11470560.
↑ Huang Q., Zhou D., Chase K., Gusella J. F., Aronin N., DiFiglia M. Immunohistochemical localization of the D1 dopamine receptor in rat brain reveals its axonal transport, pre- and postsynaptic localization, and prevalence in the basal ganglia, limbic system, and thalamic reticular nucleus (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1992. — Vol. 89, no. 24. — P. 11988—11992. — PMID 1281547.
↑ Meador-Woodruff J. H., Mansour A., Bunzow J. R., Van Tol H. H., Watson SJ Jr, Civelli O. Distribution of D2 dopamine receptor mRNA in rat brain (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1989. — Vol. 86, no. 19. — P. 7625—7628. — PMID 2529545.
↑ Le Moine C., Normand E., Guitteny A. F., Fouque B., Teoule R., Bloch B. Dopamine receptor gene expression by enkephalin neurons in rat forebrain (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1990. — Vol. 87, no. 1. — P. 230—234. — PMID 2296581.
↑ Bouthenet M. L., Souil E., Martres M. P., Sokoloff P., Giros B., Schwartz J. C. Localization of dopamine D3 receptor mRNA in the rat brain using in situ hybridization histochemistry: comparison with dopamine D2 receptor mRNA (англ.) // Brain Res. (англ.)русск. : journal. — 1991. — Vol. 564, no. 2. — P. 203—219. — DOI:10.1016/0006-8993(91)91456-B. — PMID 1839781.
↑ Le Moine C., Bloch B. Expression of the D3 dopamine receptor in peptidergic neurons of the nucleus accumbens: comparison with the D1 and D2 dopamine receptors (англ.) // Neuroscience (англ.)русск. : journal. — Elsevier, 1996. — Vol. 73, no. 1. — P. 131—143. — DOI:10.1016/0306-4522(96)00029-2. — PMID 8783237.
↑ Lévesque D., Diaz J., Pilon C., Martres M. P., Giros B., Souil E., Schott D., Morgat J. L., Schwartz J. C., Sokoloff P. Identification, characterization, and localization of the dopamine D3 receptor in rat brain using 7-3H-hydroxy-N,N-di-n-propyl-2-aminotetralin (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1992. — Vol. 89, no. 17. — P. 8155—8159. — PMID 1518841.
↑ Goldman M. E., Kebabian J. W. Aporphine enantiomers. Interactions with D-1 and D-2 dopamine receptors (англ.) // Mol. Pharmacol. (англ.)русск. : journal. — 1984. — Vol. 25, no. 1. — P. 18—23. — PMID 6231468.
↑ Cohen A. I., Todd R. D., Harmon S., O’Malley K. L. Photoreceptors of mouse retinas possess D4 receptors coupled to adenylate cyclase (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1992. — Vol. 89, no. 24. — P. 12093—12097. — PMID 1334557.
↑ Titeler M., Van Loon G. R., Seeman P., Brown G. M. D2- but not D3-dopamine receptors detected in the anterior pituitary (англ.) // Eur. J. Pharmacol. (англ.)русск. : journal. — 1981. — Vol. 71, no. 1. — P. 143—146. — PMID 7238583.
Дофамин — это… Что такое Дофамин?
Дофами́н (допами́н[1], DA) — нейромедиатор, вырабатываемый в мозгу людей и животных. Также гормон, вырабатываемый мозговым веществом надпочечников и другими тканями (например, почками), но в подкорку мозга из крови этот гормон почти не проникает. По химической структуре дофамин относят к катехоламинам. Дофамин является биохимическим предшественником норадреналина (и адреналина).
Нейромедиатор
Дофамин является одним из химических факторов внутреннего подкрепления (ФВП) и служит важной частью «системы поощрения» мозга, поскольку вызывает чувство удовольствия (или удовлетворения) чем влияет на процессы мотивации и обучения[2][3]. Дофамин естественным образом вырабатывается в больших количествах во время позитивного, по субъективному представлению человека, опыта — к примеру, секса, приёма вкусной пищи, приятных телесных ощущений, а также стимуляторов, ассоциированных с ними[4][5]. Нейробиологические эксперименты показали, что даже воспоминания о позитивном поощрении могут увеличить уровень дофамина[6][2][3][7], поэтому данный нейромедиатор используется мозгом для оценки и мотивации, закрепляя важные для выживания и продолжения рода действия[8].
Дофамин играет немаловажную роль в обеспечении когнитивной деятельности. Активация дофаминергической передачи необходима при процессах переключения внимания человека с одного этапа когнитивной деятельности на другой. Таким образом, недостаточность дофаминергической передачи приводит к повышенной инертности больного, которая клинически проявляется замедленностью когнитивных процессов (брадифрения) и персеверациями. Данные нарушения являются наиболее типичными когнитивными симптомами болезней с дофаминергической недостаточностью — например, болезни Паркинсона[9].
Как и у большинства нейромедиаторов, у дофамина существуют синтетические аналоги, а также стимуляторы его выделения в мозге. В частности, многие наркотики увеличивают выработку и высвобождение дофамина в мозге в 5—10 раз, что позволяет людям, которые их употребляют, получать чувство удовольствия искусственным образом[10][11][12]. Так, амфетамин напрямую стимулирует выброс дофамина, воздействуя на механизм его транспортировки.[13] Другие наркотики, например, кокаин и психостимуляторы, блокируют естественные механизмы обратного захвата дофамина, увеличивая его концентрацию в синаптическом пространстве[14]. Морфий и никотин имитируют действие натуральных нейромедиаторов[14], а алкоголь блокирует действие антагонистов дофамина[15]. Если пациент продолжает перестимулировать свою «систему поощрения», постепенно мозг адаптируется к искусственно повышаемому уровню дофамина, производя меньше гормона и снижая количество рецепторов в «системе поощрения»[16], что побуждает наркомана увеличивать дозу для получения прежнего эффекта. Дальнейшее развитие химической толерантности может постепенно привести к очень тяжёлым изменениям в нейронах и других структурах мозга, а в долговременной перспективе потенциально нанести серьёзный ущерб здоровью мозга[17].
Для лечения болезни Паркинсона часто используют агонисты дофаминовых рецепторов (прамипексол, бромокриптин, перголид и др.): на сегодняшний день это самая многочисленная группа противопаркинсонических средств[18]. Некоторые из антидепрессантов также обладают дофаминергической активностью[19].
Существуют и лекарственные препараты, блокирующие дофаминергическую передачу, например такие антипсихотические средства, как аминазин, галоперидол, рисперидон, клозапин и др. Резерпин блокирует накачку дофамина в пресинаптические везикулы.
При таких психических заболеваниях, как шизофрения и обсессивно-компульсивное расстройство, отмечается повышенная дофаминергическая активность в некоторых структурах мозга (при шизофрении отмечается вдобавок пониженная активность дофамина в мезокортикальном дофаминовом пути[20][21][22][23] и префронтальной коре[20][24][23]), а паркинсонизм связан с пониженным содержанием дофамина в нигростриарном пути[24][25][26][27]. Со снижением уровня дофамина в подкорковых образованиях и передних отделах головного мозга связывают также процесс нормального старения[28].
Биосинтез
Предшественником дофамина является L-тирозин (он синтезируется из фенилаланина), который гидроксилируется ферментом тирозингидроксилазой с образованием L-ДОФА, которая, в свою очередь, декарбоксилируется с помощью фермента L-ДОФА-декарбоксилазы и превращается в дофамин. Этот процесс происходит в цитоплазме нейрона.
Рецепторы
Постсинаптические дофаминовые рецепторы относятся к семейству GPCR. Существует по меньшей мере пять различных подтипов дофаминовых рецепторов — D1—5. Рецепторы D1 и D5 обладают довольно значительной гомологией и сопряжены с белком GS, который стимулирует аденилатциклазу, вследствие чего их обычно рассматривают совместно как D-1-подобные рецепторы. Остальные рецепторы подсемейства подобны D2 и сопряжены с Gi-белком, который ингибирует аденилатциклазу, вследствие чего их объединяют под общим названием D-2-подобные рецепторы. Таким образом, дофаминовые рецепторы играют роль модуляторов долговременной потенциации[29].
Участие во «внутреннем подкреплении» принимают D2 и D4 рецепторы.
В больших концентрациях дофамин также стимулирует α- и β-адренорецепторы. Влияние на адренорецепторы связано не столько с прямой стимуляцией адренорецепторов, сколько со способностью дофамина высвобождать норадреналин из гранулярных пресинаптических депо, то есть оказывать непрямое адреномиметическое действие.
«Круговорот» дофамина
Основные элементы синапса
Синтезированный нейроном дофамин накапливается в дофаминовых везикулах (т. н. «синаптическом пузырьке»). Этот процесс является протон-сопряжённым транспортом. В везикулу с помощью протон-зависимой АТФазы закачиваются ионы H+. При выходе протонов по градиенту в везикулу поступают молекулы дофамина.
Далее дофамин выводится в синаптическую щель. Часть его участвует в передаче нервного импульса, воздействуя на клеточные D-рецепторы постсинаптической мембраны, а часть возвращается в пресинаптический нейрон с помощью обратного захвата. Ауторегуляция выхода дофамина обеспечивается D2 и D3 рецепторами на мембране пресинаптического нейрона. Обратный захват производится транспортером дофамина. Вернувшийся в клетку медиатор расщепляется с помощью моноаминооксидазы (МАО) и, далее, альдегиддегидрогеназы и катехол-О-метил-трансферазы до гомованилиновой кислоты.
Участие в системе поощрения
Лабораторная крыса в специальном ящике нажимает рычаг. К голове животного прикреплены стимуляторы.
В фундаментальном исследовании 1954 года канадские учёные Джеймс Олдс и его коллега Питер Милнер обнаружили, что если имплантировать электроды в определённые участки мозга, особенно в средний узел переднего мозга, то крысу можно приучить нажимать рычаг в клетке, включающий стимуляцию низковольтными разрядами электричества[30]. Когда крысы научились стимулировать этот участок, они нажимали рычаг до тысячи раз в час[30]. Это дало основание предположить, что стимулируется центр наслаждения. Один из главных путей передачи нервных импульсов в этом участке мозга — дофаминовый, поэтому исследователи выдвинули версию, что главное химическое вещество, связанное с удовольствием, — это дофамин. В дальнейшем это предположение было подтверждено радионуклидными томографическими сканерами и открытием антипсихотиков (лекарственных средств, подавляющих продуктивные симптомы шизофрении)[31].
Дофаминергическая система
Из всех нейронов ЦНС только около семи тысяч вырабатывают дофамин. Известно несколько дофаминовых ядер, расположенных в мозге. Это дугообразное ядро (лат. nucleus arcuatum), дающее свои отростки в срединное возвышение гипоталамуса. Дофаминовые нейроны чёрной субстанции посылают аксоны в стриатум (хвостатое и чечевицеобразное ядро). Нейроны, находящиеся в области вентральной покрышки, дают проекции к лимбическим структурам и коре.
Основные дофаминовые пути.
Основными дофаминовыми путями являются:
Тела нейронов нигростриатного, мезокортикального и мезолимбического трактов образуют комплекс нейронов чёрной субстанции (англ.)русск. и вентрального поля покрышки. Аксоны этих нейронов идут вначале в составе одного крупного тракта (медиального пучка переднего мозга), а далее расходятся в различные мозговые структуры. Некоторые авторы объединяют мезокортикальную и мезолимбическую подсистемы в единую систему, однако более обоснованно выделение мезокортикальной и мезолимбической подсистем соответственно проекциям в лобную кору и лимбические структуры мозга[32].
В экстрапирамидной системе дофамин играет роль стимулирующего нейромедиатора, способствующего повышению двигательной активности, уменьшению двигательной заторможенности и скованности, снижению гипертонуса мышц. Физиологическими антагонистами дофамина в экстрапирамидной системе являются ацетилхолин и ГАМК.
Другие подсистемы
Выделяют также тубероинфундибулярный путь (лимбическая система — гипоталамус — гипофиз), инцертогипоталамический, диенцефалоспинальный и ретинальный[32] (иногда, вдобавок к этому, перивентрикулярную и ольфакторную системы[33]). Данная дифференциация не является абсолютной, поскольку проекции дофаминергических нейронов разных трактов «перекрываются»; кроме того, в мозге отмечается и диффузное распределение дофаминергических элементов (отдельных клеток с отростками)[32].
В гипоталамусе и гипофизе дофамин играет роль естественного тормозного нейромедиатора, угнетающего секрецию ряда гормонов. При этом угнетающее действие на секрецию разных гормонов реализуется при разных концентрациях дофамина, что обеспечивает высокую специфичность регуляции. Наиболее чувствительна к тормозящему действию дофаминергических сигналов секреция пролактина, в меньшей степени — секреция соматолиберина и соматотропина, в ещё меньшей — секреция кортиколиберина и кортикотропина и в совсем малой степени — секреция тиролиберина и тиротропина. Секреция гонадотропинов и гонадолиберина не угнетается дофаминергическими сигналами.
Ввиду чувствительности некоторых гормональных подсистем к уровню дофамина препараты-дофаминомиметики, усиливающие его синтез, могут применяться в качестве терапии при гормональных заболеваниях. Например, дофаминомиметики назначают при гиперпролактинемии и при болезни Паркинсона.
Дофамин и другие нейромедиаторы
Дофаминергические подсистемы находятся под контролем или сами контролируют норадренергические, серотонинергические, ГАМК-ергические, холинергические, мелатонинергические, глутаматергические, пептидергические системы. ГАМК-ергические и серотонинергические системы находятся в антагонистических отношениях с дофаминергической системой, а норадренергическая и дофаминергическая системы в различных функциональных состояниях функционируют однонаправленно: как в период бодрствования, так и в период сна. Взаимодействия дофаминергической и холинергической системы сложны, в условиях патологических процессов активность этих систем неоднозначна[33].
Гормон
Дофамин обладает рядом физиологических свойств, характерных для адренергических веществ.
Дофамин вызывает повышение сопротивления периферических сосудов (менее сильное, чем под влиянием норадреналина). Он повышает систолическое артериальное давление в результате стимуляции α-адренорецепторов. Также дофамин увеличивает силу сердечных сокращений в результате стимуляции β-адренорецепторов. Увеличивается сердечный выброс. Частота сердечных сокращений увеличивается, но не так сильно, как под влиянием адреналина.
Потребность миокарда в кислороде под влиянием дофамина повышается, однако в результате увеличения коронарного кровотока обеспечивается повышенная доставка кислорода.
В результате специфического связывания с дофаминовыми рецепторами почек дофамин уменьшает сопротивление почечных сосудов, увеличивает в них кровоток и почечную фильтрацию. Наряду с этим повышается натрийурез. Происходит также расширение мезентериальных сосудов. Этим действием на почечные и мезентериальные сосуды дофамин отличается от других катехоламинов (норадреналина, адреналина и др.). Однако в больших концентрациях дофамин может вызывать сужение почечных сосудов.
Дофамин ингибирует также синтез альдостерона в коре надпочечников, понижает секрецию ренина почками, повышает секрецию простагландинов тканью почек.
Дофамин тормозит перистальтику желудка и кишечника, вызывает расслабление нижнего пищеводного сфинктера и усиливает желудочно-пищеводный и дуодено-желудочный рефлюкс. В ЦНС дофамин стимулирует хеморецепторы триггерной зоны и рвотного центра и тем самым принимает участие в осуществлении акта рвоты.
Через гематоэнцефалический барьер дофамин мало проникает, и повышение уровня дофамина в плазме крови оказывает малое влияние на функции ЦНС, за исключением действия на находящиеся вне гематоэнцефалического барьера участки, такие как триггерная зона.
Повышение уровня дофамина в плазме крови происходит при шоке, травмах, ожогах, кровопотерях, стрессовых состояниях, при различных болевых синдромах, тревоге, страхе, стрессе. Дофамин играет роль в адаптации организма к стрессовым ситуациям, травмам, кровопотерям и др.
Также уровень дофамина в крови повышается при ухудшении кровоснабжения почек или при повышенном содержании ионов натрия, а также ангиотензина или альдостерона в плазме крови. По-видимому, это происходит вследствие повышения синтеза дофамина из ДОФА в ткани почек при их ишемии или при воздействии ангиотензина и альдостерона. Вероятно, этот физиологический механизм служит для коррекции ишемии почек и для противодействия гиперальдостеронемии и гипернатриемии.
Патологии
Наиболее известными патологиями, связанными с дофамином, являются шизофрения и паркинсонизм, а также обсессивно-компульсивное расстройство. Различные независимые исследования показали, что многие лица, страдающие шизофренией, имеют повышенную дофаминергическую активность в некоторых структурах мозга[24][25][27] (и пониженную дофаминергическую активность в мезокортикальном пути[20][23][21][22] и префронтальной коре[20][23][24]). Для лечения шизофрении применяются антипсихотики (нейролептики), которые блокируют рецепторы дофамина (преимущественно D2-типа) и варьируются в степени аффинности к другим значимым нейромедиаторным рецепторам[34]. Типичные антипсихотики в основном подавляют рецепторы D2, а новые атипичные антипсихотики и некоторые из типичных воздействуют одновременно на целый ряд нейромедиаторных рецепторов: дофамина, серотонина, гистамина, ацетилхолина и других[34].
Предполагается, что снижение уровня дофамина в мезокортикальном пути связано с негативными симптомами шизофрении[20] (сглаживание аффекта, апатия, бедность речи, ангедония, уход из общества[23]), а также с когнитивными расстройствами[20] (дефициты внимания, рабочей памяти, исполнительных функций[23]). Антипсихотическое действие нейролептиков, то есть их способность редуцировать продуктивные нарушения — бред, галлюцинации, психомоторное возбуждение — связывают с угнетением дофаминергической передачи в мезолимбическом пути[22]. Нейролептики также угнетают дофаминергическую передачу и в мезокортикальном пути[35], что при длительной терапии может приводить к усилению негативных нарушений[36].
Паркинсонизм связан с пониженным содержанием дофамина в нигростриарном пути. Наблюдается при разрушении чёрной субстанции, патологии D-1-подобных рецепторов. С угнетением дофаминергической передачи в нигростриарной системе связывают и развитие экстрапирамидных побочных эффектов при приёме антипсихотиков[22]: лекарственного паркинсонизма, дистонии, акатизии, поздней дискинезии и др.
С нарушением дофаминергической системы связывают такие расстройства, как ангедония, депрессия, деменция, патологическая агрессивность, фиксация патологических влечений, синдром персистирующей лактореи-аменореи, импотенция, акромегалия, синдром беспокойных ног и периодических движений в конечностях[33].
Процесс старения
По данным исследований, процесс старения проявляется уменьшением объема и массы головного мозга и уменьшением числа синаптических связей; кроме уменьшения числа церебральных рецепторов, имеет место и медиаторная церебральная недостаточность. С возрастом уменьшается количество и плотность дофаминовых D2-рецепторов стриатума, снижается концентрация дофамина в подкорковых образованиях головного мозга. Клиническими проявлениями этих изменений являются обеднение мимики, некоторая общая замедленность, сгорбленная, старческая поза, укорочение длины шага.
«Дофамин-чувствительные» изменения отмечаются также в когнитивной сфере: с возрастом снижается быстрота реакции, становится труднее усваивать и реализовывать новую программу действия, снижается уровень внимания, объём оперативной памяти. При отсутствии органической патологии возрастные когнитивные изменения не приводят к дезадаптации пожилых людей и позволяют поддерживать привычный ритм социальной активности[28].
См. также
Примечания
↑ В русской терминологии принято название «дофамин». Вариант «допамин» представляет собой прямую транслитерацию принятого в западной литературе термина dopamine и отсутствует в русских словарях. Различие объясняется разницей в сокращениях, возникшей из-за соответствия ph = ф: dihydroxyphenylalanine = DOPA, дигидроксифенилаланин = ДОФА.
↑ 12 It’s all about dopamine. Архивировано из первоисточника 22 августа 2011.
↑ 12 Biology of Happiness. Архивировано из первоисточника 22 августа 2011.
↑ Допамин — удовольствие от жизни. Архивировано из первоисточника 3 февраля 2012.
↑ Ваш мозг во время секса. Архивировано из первоисточника 22 августа 2011.
↑ Удовольствие: дофамин. Архивировано из первоисточника 24 января 2012.
↑ Кокаиновые наркоманы могут получать удовольствие только от одного ожидания этого удовольствия. Архивировано из первоисточника 24 января 2012.
↑ Dopamine and desire. Архивировано из первоисточника 22 августа 2011.
↑Захаров В.В., Яхно Н.Н. Когнитивные расстройства в пожилом и старческом возрасте: Методическое пособие для врачей. — Москва, 2005.
↑ Мозг и наркотики. Архивировано из первоисточника 22 августа 2011.
↑ Your Brain on Drugs: Dopamine and Addiction. Архивировано из первоисточника 22 августа 2011.
↑ Dopamine — A Sample Neurotransmitter. Архивировано из первоисточника 22 августа 2011.
↑ A Mechanism for Amphetamine-Induced Dopamine Overload.
↑ 12 HOW DRUGS AFFECT NEUROTRANSMITTERS. Архивировано из первоисточника 22 августа 2011.
↑ Мужчины чаще становятся алкоголиками из-за дофамина, выяснили ученые. Архивировано из первоисточника 22 августа 2011.
↑ Addiction and Dopamine (D2) Receptor Levels (2006). Архивировано из первоисточника 22 августа 2011.
↑ The Science Behind Drug Use and Addiction. Архивировано из первоисточника 22 августа 2011.
↑Катунина Е. А. Агонисты дофаминовых рецепторов и проблема эквивалентности доз // Неврология. — 2010. — № 2.
↑Shelton RC, Tomarken AJ Can Recovery From Depression Be Achieved? // American Psychiatric Association Psychiatr Serv. — November 2001. — № 52. — С. 1469—1478. Перевод: Можно ли достичь выздоровления при депрессии? // Обзор современной психиатрии. — November 2004. — В. 21.
↑ 123456 Abi-Dargham A, Moore H (October 2003). «Prefrontal DA transmission at D1 receptors and the pathology of schizophrenia». Neuroscientist9 (5): 404–16. DOI:10.1177/1073858403252674. PMID 14580124.
↑ 12 Lieberman JA (2004). «Dopamine partial agonists: a new class of antipsychotic». CNS Drugs18 (4): 251–67. PMID 15015905.
↑ 1234 Справочное руководство по психофармакологическим и противоэпилептическим препаратам, разрешенным к применению в России / Под ред. С. Н. Мосолова. — 2-е, перераб. — М.: «Издательство БИНОМ», 2004. — С. 17. — 304 с. — 7000 экз. — ISBN 5-9518-0093-5
↑ 123456Abi-Dargham A The Dopamine Hypothesis of Schizophrenia. Schizophrenia Research Forum (5 December 2005). Архивировано из первоисточника 3 февраля 2012. Проверено 26 сентября 2011. Перевод: Дофаминовая гипотеза шизофрении (15 August 2008). Архивировано из первоисточника 3 февраля 2012.
↑ 1234 The Dopamine Hypothesis of Schizophrenia: Version III—The Final Common Pathway. Архивировано из первоисточника 23 августа 2011.
↑ 12 Increased Striatal Dopamine Transmission in Schizophrenia: Confirmation in a Second Cohort. Архивировано из первоисточника 23 августа 2011.
↑ Presynaptic Regulation of Dopamine Transmission in Schizophrenia. Архивировано из первоисточника 23 августа 2011.
↑ 12 Dysconnection in Schizophrenia: From Abnormal Synaptic Plasticity to Failures of Self-monitoring. Архивировано из первоисточника 23 августа 2011.
↑ Mehta M.A., Riedel W.J. Dopaminergic Enhancement of Cognitive Function. // Curr. Pharm. Des. 2006. v. 12. pp. 2487—2500.
↑ 12 Британская энциклопедия, см. раздел «Reward and punishment». Архивировано из первоисточника 3 февраля 2012.
↑ The Functional Neuroanatomy of Pleasure and Happiness. Архивировано из первоисточника 3 февраля 2012.
↑ 123Орловская Д. Д. Нейрохимические системы мозга // Общая психиатрия / Под ред. А. С. Тиганова. — Москва, 2006.
↑ 123Левин Я.И. Нейрохимическая медицина. Часть 1. Церебральные дофаминергические системы // Современная терапия психических расстройств. — 2008. — № 1.
↑ 12 A Roadmap to Key Pharmacologic Principles in Using Antipsychotics. PubMed (USA).
↑ Basu D, Marudkar M, Khurana H (2000). «Abuse of neuroleptic drug by psychiatric patients» 54 (2): 59—62. PMID 11271726.
↑Бородин В. И. Сперидан (рисперидон) при терапии шизофренических психозов // Психиатрия и психофармакотерапия. — 2006. — В. 8 (4).
Литература
Ашмарин И. П., Ещенко Н. Д., Каразеева Е. П. Нейрохимия в таблицах и схемах. — М.: Экзамен, 2007.
Васильев В. Н. Диагностика и терапия инкурабельных нервных и психических заболеваний допаминовой этиологии. Биокоррекция Васильева. — М.: Медиакит, 2009. — 247 с. — ISBN 978-5-9901746-1-0
тверда, біла порошкоподібна з характерним запахом речовина
Густина
1,26 г/см³ [джерело?]
Тпл
128°C [джерело?]
Розчинність (вода)
60 г у 100 мл води [джерело?]
Кислотність (pKa)
8,93 [джерело?]
Якщо не зазначено інше, дані наведено для речовин у стандартному стані (за 25 °C, 100 кПа)
Інструкція з використання шаблону
Примітки картки
Дофамі́н(допамін) (4-(2-аміноетил)бензол-1,2-діол) — нейромедіатор, біологічно активна хімічна речовина, яка в мозку людини передає емоційну реакцію і дозволяє відчувати задоволення і не відчувати біль, також нейромедіатор, відповідальний за рух, тому виробляється у певних клітинах мозку, що контролюють м’язову активність.
Дофамін викликає почуття щастя й ейфорії. Також дофамін є одним з головних компонентів біохімічного механізму закоханості — мозок людини, що закохалася, починає стрімко виробляти дофамін. Підвищення концентрації дофаміну спостерігається під час споживання приємної на смак їжі чи роботи за хорошу винагороду. Вживання речовин, що викликають залежність: алкоголь, нікотин чи кокаїн, тимчасово збільшує концентрацію дофаміну в мозку. У тих, хто страждає на депресію, його рівень низький. Також нестача дофаміну призводить до сповільненості та загальмованості когнітивних процесів в мозку людини, скутості рухів, утруднення ходи, обмеженості рухової активності.
Дофамін є життєво необхідною речовиною, нестача якої зумовлює погане самопочуття (пригнічений емоційний стан), а також призводить до дуже серйозних розладів руху та м’язової активності (наприклад хвороба Паркінсона).
Окрім нейромедіаторних властивостей у центральній нервовій системі, дофамін діє як гормон. Він здійснює специфічний вплив на функцію серцево-судинної системи — підвищує систолічний артеріальний тиск в результаті стимуляції α-адренорецепторів, збільшує силу серцевих скорочень в результаті стимуляції β-адренорецепторів, збільшує серцевий викид і частоту серцевих скорочень. Також дофамін спричиняє дилатацію судин нирок, збільшує діурез та натрійурез, підвищує синтез простагландинів тканиною нирок, стимулює екзокринну функцію підшлункової залози.
Ферментативний розпад дофаміну в синаптичній щілині
Процес синтезу дофаміну відбувається в цитоплазмі нейрона. При гідроксилюванні L-тирозину (який синтезується з фенілаланіну) за допомогою тирозин-гідроксилази утворюється L-ДОФА (дигідроксифенілаланін), що за допомогою L-ДОФА-декарбоксилази перетворюється в дофамін. Під дією дофамін-β-гідроксилази дофамін може перетворюватись на норепінефрин, з якого за допомогою фенілетаноламін-N-метилтрансферази синтезується епінефрин.
Синтезований нейроном дофамін накопичується у везикулах. Цей процес називається протон-спряженим транспортом — у везикулу за допомогою протон-залежної АТФази накачуються йони Н+, і при їхньому виході за градієнтом у везикулу потрапляють молекули дофаміну. Далі дофамін виводиться в синаптичну щілину. Частина його бере участь в передачі нервового імпульсу, діючи на дофамінові рецептори постсинаптичної мембрани, а частина повертається в пресинаптичний нейрон за допомогою дофамінового транспортера. Дофамін, що повернувся в клітину, може розщеплюватись за допомогою моноамінооксидази.
Функції (дофамін як гормон)[ред. | ред. код]
Дофамін мало проникає через гематоенцефалічний бар’єр, і синтез та функції допафіну у тілі великою мірою незалежні від синтезу та функцій допаміну у головному мозку
Дофамін має низку фізіологічних властивостей, характерних для адренергічних речовин.
Вплив на серце, судини[ред. | ред. код]
Дофамін викликає підвищення опору периферичних судин (не таке сильне, як під впливом норадреналіну). Підвищує систолічний артеріальний тиск в результаті стимуляції α-адренорецепторів. Також дофамін збільшує силу серцевих скорочень в результаті стимуляції β-блокаторів. Збільшує серцевий викид. Збільшує частоту серцевих скорочень, але не так сильно, як під впливом адреналіну.
Підвищує потребу міокарду в кисні під впливом дофаміну, проте в результаті збільшення коронарного кровотоку забезпечує підвищена доставку кисню.
Вплив на нирки[ред. | ред. код]
В результаті специфічного зв’язування з дофаміновими рецепторами нирок дофамін зменшує опір ниркових судин, збільшує в них кровотік і ниркову фільтрацію. Поряд з цим підвищується натрійурез. Відбувається також розширення мезентеріальних судин. Цією дією на ниркові і мезентеріальні судини дофамін відрізняється від інших катехоламінів (норадреналіну, адреналіну і т.д.). Однак у великих концентраціях дофамін може викликати звуження ниркових судин.
Дофамін також інгібує синтез альдостерону в корі надниркових залоз, знижує секрецію реніну нирками, підвищує секрецію простагландинів тканиною нирок.
Вплив на травлення[ред. | ред. код]
Дофамін гальмує перистальтику шлунка і кишечника, викликає розслаблення нижнього стравохідного сфінктера і підсилює шлунково-стравохідний і дуодено-шлунковий рефлюкс. В ЦНС дофамін стимулює хеморецептори триггерної зони і блювотного центру і тим самим бере участь у здійсненні акту блювоти.
Вплив на нервову систему[ред. | ред. код]
Через гематоенцефалічний бар’єр дофамін мало проникає, і підвищення рівня дофаміну в плазмі крові мало впливає на функції ЦНС, за винятком дії на ділянки, що знаходяться поза гематоенцефалічним бар’єром, такі як тригерна зона.
Підвищення рівня дофаміну[ред. | ред. код]
Підвищення рівня дофаміну в плазмі крові відбувається під час шоку, травм, опіків, крововтраті, стресових станах, при різних больових синдромах, тривозі, страху. Дофамін грає роль в адаптації організму до стресових ситуацій, травм, крововтраті і т.д.
Рівень дофаміну в крові так само підвищується при погіршенні кровопостачання нирок або при підвищеному вмісті іонів натрію, а також ангіотензиногену чи альдостерону в плазмі крові. Імовірно, це відбувається внаслідок підвищення синтезу дофаміну з ДОФА в тканини нирок при їх ішемії або при впливі ангіотензиногену і альдостерону. Ймовірно, цей фізіологічний механізм служить для корекції ішемії нирок та для протидії гіперальдостеронемії і гіпернатріємії.
Обсуждение:Дофамин — Википедия
Материал из Википедии — свободной энциклопедии
Как лучше разнести дофамин (лекарство) и дофамин (нейромедиатор)? Или оставить так (тоже касается остальных медиаторов использующихся как лекарство).
Именно так и разнести, дофамин (гормон) и дофамин (лекарственный препарат). Или, если в виде лекарства он используется, скажем, в виде гидрохлорида или гидротартрата, то соответственно просто дофамин и дофамина гидрохлорид… Rombik 12:00, 23 августа 2005 (UTC)
дофамин как гормон не смотрела, вдруг там че не так, не знаю тоже. —Ликка 16:10, 23 декабря 2008 (UTC)
В статье говорилось, что шизофрения связана с ПОВЫШЕНИЕМ дофамина в мезокортикальном пути. О мезолимбическом не говорилось ни слова.
Подправила. Кроме того, внесла и другие изменения.
Пожалуйста, будьте внимательней при написании.
V for Vendetta 15:59, 7 января 2009 (UTC) V for Vendetta
ну, увы, патологии — не моя специальность… я извиняюсь, но тут дело не во внимательности, а так сказать, в недостаточной компетенции в шизофрении. надо было просто как-то обозначить раздел для дальнейшей дораьотки. какбэ… правим смело. спасибо. —Ликка 12:11, 8 января 2009 (UTC)
Допамин — удовольствие от жизни[править код]
Допамин — удовольствие от жизни В мозгу млекопитающих существует так называемый «центр удовольствия», состоящий из пучка нейронов. Все наши удовольствия, независимо от причины их происхождения — сопровождаются выбросом допамина…
Возможно, у центре удовольствия и допамине следует написать что-то в статье?
Seleonov 19:57, 6 ноября 2010 (UTC)
Во-первых, центр удовольствий является не более чем предположением, во-вторых, нет доказанной связи между удовольствиями и допамином.
«Вариант „допамин“ употребляется только в специальной литературе, так как является калькой с английского dopamine»
Wiki:»Словообразовательные кальки — это слова, полученные «поморфемным» переводом иностранного слова на русский язык.»
Очевидно, «допамин» калькой не является. Это слово — недоразумение. Dihydroxyphenyl — Дигидроксифенил.
С уважением
109.239.36.117 11:24, 8 августа 2011 (UTC)Кирилл
Это гормон не удовольствия[править код]
Это гормон не удовольствия, а гормон «обещания удовольствия».
Вот в этой книге написано http://www.ozon.ru/context/detail/id/8243294/
Поищите может где скачать можно или выдержки посмотреть. Там есть ссылки на тех кто это доказал.
А то устаревшие данные в статье получаются.
 Основные элементы синапса
Основные элементы синапса Лабораторная крыса в специальном ящике нажимает рычаг. К голове животного прикреплены стимуляторы.
Лабораторная крыса в специальном ящике нажимает рычаг. К голове животного прикреплены стимуляторы.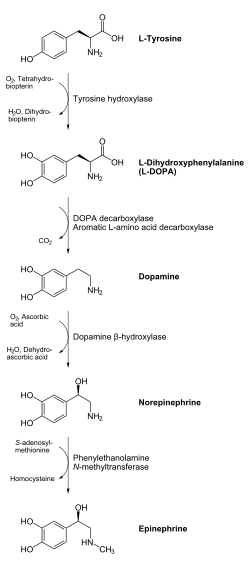
 Основные элементы синапса
Основные элементы синапса Лабораторная крыса в специальном ящике нажимает рычаг. К голове животного прикреплены стимуляторы.
Лабораторная крыса в специальном ящике нажимает рычаг. К голове животного прикреплены стимуляторы.
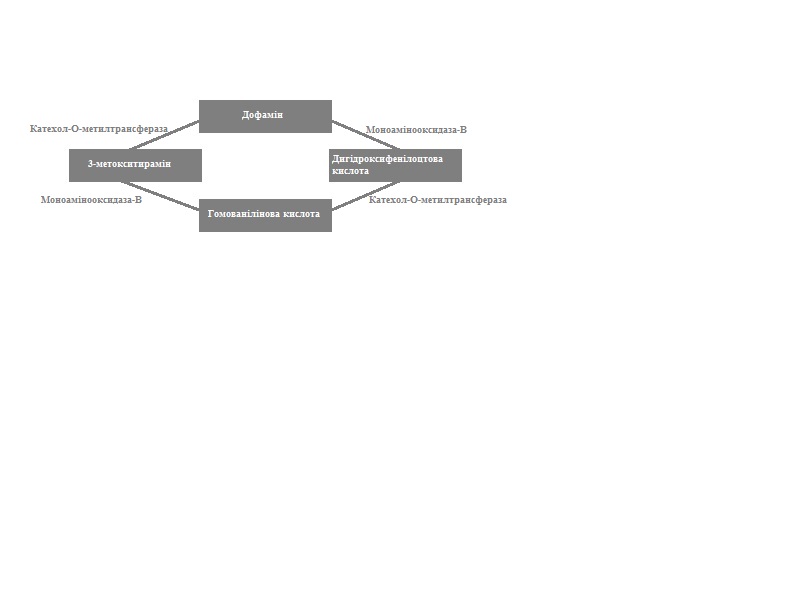
 Ферментативний розпад дофаміну в синаптичній щілині
Ферментативний розпад дофаміну в синаптичній щілині
Добавить комментарий