Где синтезируются углеводы
7.3. Биосинтез углеводов
Впроцессе метаболизма углеводов происходит не только их катаболизм, но и взаимопревращение углеводов, биосинтез углеводов из неуглеводных предшественников и синтез полисахаридов из моносахаридов.
Общим центральным путем биосинтеза углеводов из неуглеводных предшественников является синтез глюкозы из пировиноградной кислоты, реализующийся путём обращения большинства реакций гликолиза . При этом для трех практически необратимых реакций(гексокиназной реакции, фосфофруктокиназной реакции и пируваткиназной реакции) существуют обходные ферментативные пути, обеспечивающие обратное превращение. В частности, внутри митохондрий пировиноградная кислота не может превратиться фосвфоенолпировиноградную кислоту путём обращения пируваткиназной реакции,
ипоэтому в начале под влиянием митохондриальной пируваткарбоксилазы пировиноградная кислота карбоксилируется в щавелево-уксусную кислоту. Реакция идёт при участии АТФ и положительного модулятора( ктиватора) фермента пируваткарбоксилазы – ацетил-КоА. Пируваткарбоксилаза является регуляторным ферментом. Реакция протекает в митохондриях.
Ch5 | COOH | ||||
пируваткарбоксилаза | |||||
CO + CO + H O + АТФ | Ch4 | + АДФ +Ф | |||
2 | ацетил-КоА | ||||
COOH | CO |
пировиноградная
кислота | COOH | |
щавелевоуксусная | ||
кислота (оксалоацетат) |
Оксалоацетат восстанавливается затем в митохондриях в малат:
COOH | митохондриальная | COOH | |||||
CH | малатдегидрогеназа | Ch4 | +НАД | ||||
2 | + НАД.Н 2 | CHOH | |||||
CO | |||||||
COOH | COOH | ||||||
оксалоацетат | малат |
(яблочная кислота)
Малат переходит из митохондрий в цитоплазму (с помощью пермиаз), где окисляется цитоплазматической малатдегидрогеназой до оксалоацетата:
COOH | цитоплазматическая | COOH | ||||
Ch4 | малатдегидрогеназа | CH | ||||
CHOH | +НАД | 2 | + НАД.Н 2 | |||
CO | ||||||
COOH | COOH | |||||
малат | оксалоацетат |
Под действием фосфоенолпируваткарбоксикиназы (фосфопируваткарбоксилазы) далее происходит превращение оксалоацетата в фосфоенолпируват. При этом донором фосфата служит гуанозинтрифосфат(ГТФ) или инозинтрифосфат (ИТФ):
COOH | фосфоенолпируват- | Ch4 | ||||||||
карбоксикиназа | OH | |||||||||
Ch4 | Mg++ | |||||||||
+ ГТФ | C | O | P | O + CO 2 + ГДФ | ||||||
CO | ||||||||||
COOH | OH | |||||||||
фосфоенолпируват | ||||||||||
COOH |
оксалоацетат
Далее фосфоенолпируват в силу обратимости реакций гликолиза легко превращается в фруктозо-1,6-дифосфат. Обратное превращение фруктозо-1,6- дифосфата в фруктозо-6-фосфат и далее глюкозо-6-фосфата в глюкозу обеспечивается соответствующими специфическими фосфатазами – дифосфофруктозофосфатазой и глюкозо-6-фосфатазой. Первая из них является регуляторным ферментом. Субстратами для синтеза глюкозы могут служить не только пируват, но и все вещества неуглеводной природы, способные превращаться в пируват или в один из метаболитов гликолиза или оксалоацетат. К таким веществам относится глицерин, который через α-глицерофосфат и фосфодиоксиацетон способен превращаться в фосфоглицериновый альдегид, т.е. в метаболит гликолиза, а также лактат. К этим веществам относятся и промежуточные продукты лимоннокислого цикла постольку, поскольку они могут превращаться в оксалоацетат. Однако главным источником новообразования глюкозы являются аминокислоты, превращающиеся в пируват и в оксалоацетат. Синтез глюкозы из неуглеводных источников получил название глюконеогенеза.
Ни ацетил-КоА, ни СО2 не могут превращаться в глюкозу в тканях животных. Кроме того, в животных тканях ацетил-КоА не может непосредственно превращаться ни в пируват, ни в сукцинат. У высших животных отсутствует метаболический путь превращения атомов углерода жирных кислот в углеводы. В отличии от этого, растения и многие микроорганизмы могут синтези-
ровать углеводы из жирных кислот через ацетил-КоА при помощи реакций глиоксилатного цикла.
В процессе метаболизма углеводов происходит перестройка одного моносахарида в другой. Причем, многие биологически важные сахара могут образовываться не только в процесса апотомического распада углеводов, но и другим путем. Особенно реакционно-способными формами моносахаридов, наряду с их фосфорными эфирами, являются их соединения с пиримидиновыми нуклеотидами, в частности, с уридиндифосфатом. Моносахариды в соединении с уридиндифосфатом легко подвергаются изомеризации и гликозилтрансферазным реакциям. При участии уридиндифосфата осуществляются синтезы
дисахаридов. Уридиндифосфат принимает также участие в синтезе гликогена из глюкозы.
Как уже говорилось ранее, практически вся всосавшаяся из пищеварительного тракта глюкоза поступает в печень, где быстро при участии АТФ подвергается реакции фосфорилирования, катализируемой гексокиназой и далее
Сайт учителей биологии МБОУ Лицей № 2 города Воронежа
Биосинтез углеводов — фотосинтез
Биосинтез белка создает полимерную молекулу из готовых мономеров – аминокислот, уже имеющихся в клетке. Этот процесс осуществляется за счет внутренней энергии клетки (АТФ).
Биосинтез углеводов идет принципиально иначе. В клетках растений мономеры – моносахариды – образуются из неорганических веществ (углекислого газа и воды). Осуществляется этот процесс с помощью энергии света, поступающей в клетку из внешней среды. Этот процесс называют фотосинтезом (от греч. photos – «свет» и synthesis – «соединение»).
Созданные в клетке моносахариды (глюкоза, фруктоза) как первичные продукты фотосинтеза используются затем для биосинтеза различных полисахаридов, сложных белковых соединений, жирных кислот, нуклеиновых кислот и многих других органических соединений.
Фотосинтез – процесс, чрезвычайно важный для всего живого населения планеты. Он происходит в клетках зеленых растений с помощью пигментов (
Хлоропласты – это внутриклеточные органоиды (пластиды), которые благодаря пигменту хлорофиллу окрашены в зеленый цвет. В растительной клетке обычно содержится от 15 до 50 хлоропластов.
Фотосинтез – сложный многоступенчатый процесс. Начало ему задает свет. Многолетние исследования фотосинтеза показали, что он включает в себя две стадии: световую и темновую.
Первая стадия фотосинтеза – световая. Под действием энергии света молекулы хлорофилла (и других соединений, называемых переносчиками) возбуждаются и теряют электроны. Часть электронов, захваченных ферментами, способствует образованию АТФ путем присоединения остатка фосфорной кислоты (Ф) к АДФ. Другая часть электронов принимает участие в расщеплении (разложении) воды на молекулярный кислород, ионы водорода и электроны. Разложение воды происходит внутри хлоропласта.
Образовавшийся при расщеплении воды водород с помощью электронов присоединяется к веществу, способному транспортировать водород в пределах хлоропласта. Таким веществом является сложное органическое соединение из группы ферментов – окисленный никотинамидаденин-динуклеотидфосфат, или НАДФ. Присоединив водород, НАДФ восстанавливается до НАДФ • Н. В такой химической связи запасается энергия, и заканчивается первая стадия фотосинтеза.
Участие энергии света здесь является обязательным условием. Поэтому данную стадию называют еще стадией световых реакций.
Кислород, образующийся на первой стадии фотосинтеза как побочный продукт при расщеплении воды, выводится наружу или используется клеткой для дыхания.
Вторая стадия фотосинтеза – темновая. Здесь используются образовавшиеся в процессе световых реакций продукты. С их
помощью происходит преобразование углекислого газа в простые углеводы – моносахариды.
Их создание идет путем большого количества реакций восстановления СО
Все световые реакции (первая стадия фотосинтеза) происходят на мембранах хлоропласта – в тилакоидах, а темновые (вторая стадия фотосинтеза) – между мембранами внутри хлоропласта – в строме.
Сложный поэтапный процесс фотосинтеза идет непрерывно, пока зеленые клетки получают световую энергию.
На скорость фотосинтеза влияют внешние условия среды: интенсивность освещения, концентрация углекислого газа и температура. Если эти параметры достигают оптимальных величин, происходит усиление фотосинтеза. Благодаря фотосинтезу примерно 1–1,5% энергии Солнца, получаемой зелеными растениями, запасается в органических молекулах. Фотосинтезирующие организмы дают пищу гетеротрофам, а также кислород, необходимый для дыхания всем живым существам на планете. Установлено, что 21% кислорода в современной атмосфере Земли создан главным образом путем фотосинтеза.
Фотосинтез – уникальный процесс создания зелеными клетками органических веществ из неорганических, притом идущий в огромных масштабах на суше и в воде. Ежегодно растения связывают 1,7 млрд т углерода, образуя при этом более 150 млрд т органического вещества и выделяя около 200 млрд т кислорода.
Фотосинтез – единственный на нашей планете процесс превращения энергии солнечного света в энергию химических связей органических веществ. Таким способом энергия Солнца, поступившая из космоса, преобразуется и запасается клетками зеленых растений в углеводах, белках и липидах, обеспечивая жизнедеятельность всего гетеротрофного населения живого мира – от бактерий до человека.
Вот почему выдающийся русский ученый–естествоиспытатель К. А. Тимирязев эту роль зеленых растений для жизни на Земле назвал космической.
< Предыдущая страница «Биосинтез белков»
Следующая страница «Энергетический обмен» >
7.3. Биосинтез углеводов

196 | 7. Обмен углеводов |
Впроцессе метаболизма углеводов происходит не только их катаболизм, но и взаимопревращение углеводов, биосинтез углеводов из неуглеводных предшественников и синтез полисахаридов из моносахаридов.
Общим центральным путем биосинтеза углеводов из неуглеводных предшественников является синтез глюкозы из пировиноградной кислоты, реализующийся путём обращения большинства реакций гликолиза . При этом для трех практически необратимых реакций(гексокиназной реакции, фосфофруктокиназной реакции и пируваткиназной реакции) существуют обходные ферментативные пути, обеспечивающие обратное превращение. В частности, внутри митохондрий пировиноградная кислота не может превратиться фосвфоенолпировиноградную кислоту путём обращения пируваткиназной реакции,
ипоэтому в начале под влиянием митохондриальной пируваткарбоксилазы пировиноградная кислота карбоксилируется в щавелево-уксусную кислоту. Реакция идёт при участии АТФ и положительного модулятора( ктиватора) фермента пируваткарбоксилазы – ацетил-КоА. Пируваткарбоксилаза является регуляторным ферментом. Реакция протекает в митохондриях.
Ch4 |
| COOH |
| ||
|
| пируваткарбоксилаза |
|
| |
| CO + CO + H O + АТФ |
|
| Ch3 | + АДФ +Ф |
|
|
| |||
| 2 | ацетил-КоА |
|
| |
| COOH |
|
| CO |
|
пировиноградная
кислота | COOH | |
щавелевоуксусная | ||
| ||
| кислота (оксалоацетат) |
Оксалоацетат восстанавливается затем в митохондриях в малат:
| COOH |
| митохондриальная |
|
| COOH |
|
|
|
|
|
| |||
| CH |
| малатдегидрогеназа |
| Ch3 | +НАД | |
| 2 | + НАД.Н 2 |
|
|
| CHOH | |
CO |
|
|
| ||||
| COOH |
|
|
|
| COOH |
|
|
|
|
|
| |||
оксалоацетат |
|
| малат |
| |||
(яблочная кислота)
Малат переходит из митохондрий в цитоплазму (с помощью пермиаз), где окисляется цитоплазматической малатдегидрогеназой до оксалоацетата:
| COOH |
| цитоплазматическая | COOH |
| |
|
|
|
|
|
| |
| Ch3 |
| малатдегидрогеназа |
| CH |
|
| CHOH | +НАД |
|
| 2 | + НАД.Н 2 |
|
| CO | ||||
| COOH |
|
|
| COOH |
|
|
|
|
|
| ||
малат |
|
| оксалоацетат | |||
7. Обмен углеводов | 197 |
Под действием фосфоенолпируваткарбоксикиназы (фосфопируваткарбоксилазы) далее происходит превращение оксалоацетата в фосфоенолпируват. При этом донором фосфата служит гуанозинтрифосфат(ГТФ) или инозинтрифосфат (ИТФ):
COOH |
| фосфоенолпируват- |
| Ch3 |
|
|
| ||||
| карбоксикиназа |
|
| OH | |||||||
|
|
|
|
| |||||||
Ch3 |
| Mg++ |
|
|
|
|
|
|
|
| |
+ ГТФ | C |
| O | P |
| O + CO 2 + ГДФ | |||||
|
| ||||||||||
|
|
|
|
| |||||||
|
|
|
|
|
|
| |||||
| CO |
| |||||||||
|
|
| COOH |
| OH | ||||||
|
|
|
| фосфоенолпируват | |||||||
COOH |
|
| |||||||||
оксалоацетат
Далее фосфоенолпируват в силу обратимости реакций гликолиза легко превращается в фруктозо-1,6-дифосфат. Обратное превращение фруктозо-1,6- дифосфата в фруктозо-6-фосфат и далее глюкозо-6-фосфата в глюкозу обеспечивается соответствующими специфическими фосфатазами – дифосфофруктозофосфатазой и глюкозо-6-фосфатазой. Первая из них является регуляторным ферментом. Субстратами для синтеза глюкозы могут служить не только пируват, но и все вещества неуглеводной природы, способные превращаться в пируват или в один из метаболитов гликолиза или оксалоацетат. К таким веществам относится глицерин, который через α-глицерофосфат и фосфодиоксиацетон способен превращаться в фосфоглицериновый альдегид, т.е. в метаболит гликолиза, а также лактат. К этим веществам относятся и промежуточные продукты лимоннокислого цикла постольку, поскольку они могут превращаться в оксалоацетат. Однако главным источником новообразования глюкозы являются аминокислоты, превращающиеся в пируват и в оксалоацетат. Синтез глюкозы из неуглеводных источников получил название глюконеогенеза.
Ни ацетил-КоА, ни СО2 не могут превращаться в глюкозу в тканях животных. Кроме того, в животных тканях ацетил-КоА не может непосредственно превращаться ни в пируват, ни в сукцинат. У высших животных отсутствует метаболический путь превращения атомов углерода жирных кислот в углеводы. В отличии от этого, растения и многие микроорганизмы могут синтези-
ровать углеводы из жирных кислот через ацетил-КоА при помощи реакций глиоксилатного цикла.
В процессе метаболизма углеводов происходит перестройка одного моносахарида в другой. Причем, многие биологически важные сахара могут образовываться не только в процесса апотомического распада углеводов, но и другим путем. Особенно реакционно-способными формами моносахаридов, наряду с их фосфорными эфирами, являются их соединения с пиримидиновыми нуклеотидами, в частности, с уридиндифосфатом. Моносахариды в соединении с уридиндифосфатом легко подвергаются изомеризации и гликозилтрансферазным реакциям. При участии уридиндифосфата осуществляются синтезы

198 | 7. Обмен углеводов |
дисахаридов. Уридиндифосфат принимает также участие в синтезе гликогена из глюкозы.
Как уже говорилось ранее, практически вся всосавшаяся из пищеварительного тракта глюкоза поступает в печень, где быстро при участии АТФ подвергается реакции фосфорилирования, катализируемой гексокиназой и далее может не только распадаться, но и участвовать в синтезе гликогена. В этом случае образовавшийся глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фосфат. Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена.
Этот синтез, согласно Лелуару, осуществляется следующим путем. Глю- козо-1-фосфат в присутствии особого фермента(гликозилтрансферазы) вступает во взаимодействие с уридинтрифосфатом (УТФ) с образованием уридиндифосфоглюкозы (УДФ-Глюк.) и пирофосфата. Последний под влиянием пирофосфатазы немедленно подвергается гидролизу.
гексокиназа + АТФ
АДФ
|
|
|
|
|
|
| OH |
|
|
| |
|
| Ch3 | O P |
| O |
|
|
| |||
|
|
|
|
|
| ||||||
|
|
|
|
|
| ||||||
H |
| O |
|
| OH |
|
|
| |||
| H | фосфоглюкомутаза | |||||||||
| H |
|
|
|
|
|
| ||||
| OH | H |
|
|
|
|
|
|
| ||
|
|
|
|
|
|
| |||||
HO |
|
|
|
|
|
|
|
|
|
| |
|
| OH |
|
|
| ||||||
|
|
|
|
|
|
|
|
|
| ||
| H | OH |
|
|
| ||||||
| глюкоза |
|
| глюкозо-6-фосфат |
| |||||
|
| Ch3OH |
|
|
|
|
| |||
H |
| O H |
|
| гликозилтрансфераза + УТФ |
| ||||
| H |
|
|
|
| OH | УДФГ | |||
| OH | H |
|
|
|
| ||||
HO |
|
|
|
|
| P O |
|
| уридиндифосфоглюкоза | |
| O |
|
| ФФ | ||||||
|
|
|
| |||||||
| H |
| OH |
| OH |
|
|
| ||
глюкозо-1-фосфат | Ф | Ф |
| |||||||
Далее в присутствии особой трансферазы, получившей название гликогенсинтетазы и «затравочного» количества гликогена (полиглюкозы, имеющей не менее четырех остатков глюкозы) происходит своеобразный ферментативный процесс – удлинение цепочки гликогена с невосстанавливающего конца с образованием 1-4 связи за счет присоединения остатков глюкозы, входящих в состав УДФ-глюкозы.

|
|
|
|
|
|
|
|
|
|
|
| 7. Обмен углеводов |
|
|
|
|
|
|
|
|
| 199 | |||||
|
|
|
|
| Ch3OH |
|
|
|
|
|
|
|
|
|
|
| Ch3OH |
|
|
|
| ||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| ||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| H |
| O H |
|
|
|
| ||||||
|
|
| H |
| O | H |
|
|
| гликогенсинтетаза |
|
|
|
|
|
|
|
| |||||||||
|
|
|
|
|
|
|
| +УДФГ |
|
|
|
| H |
|
|
|
|
|
| + УДФ | |||||||
|
|
|
| H |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| OH | H |
|
|
|
| |||||||
|
|
|
| OH | H |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| ||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
| O |
|
|
|
|
|
|
|
| |||||
|
|
| O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| O |
|
|
|
| ||||
|
|
|
|
|
| O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| H |
| OH |
|
|
| n+1 | ||
|
|
|
| H | OH |
|
| n | |||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |||||||||
полиглюкоза
Образовавшийся УДФ затем вновь фосфорилируется за счет АТФ до УТФ и, таким образом, весь цикл превращения глюкозо-1-фосфата может начинаться сначала. Ветвление синтезируемой молекулы гликогена, как было показано А.Н. Петровой, осуществляется при участии особого фермента, катализирующего замыкание связей между 1 и 6 углеродными атомами остатков глюкозыамилотрансгликозилазы.
7.4. Нейрогуморальная регуляция углеводного обмена. Роль печени в углеводном обмене
Существенная роль в углеводном обмене принадлежит печени. Синтезируемый гликоген с его огромным молекулярным весом (от нескольких сот до нескольких миллионов) может накапливаться в печени в значительных количествах (у человека около 150 г) без изменения осмотических условий в клетках, и, следовательно, является удобной формой депонирования глюкозы. Повидимому, в этой связи печень играет важнейшую роль в поддержании постоянства содержания глюкозы в крови.
Наряду с печенью, значительное место в регуляции обмена углеводов принадлежит периферическим органам и прежде всего мышцам, усиленно потребляющим глюкозу. Однако в первую очередь печень сглаживает колебания уровня сахара, возникающие из-за неравномерного поступления его в кровь. При повышении концентрации сахара в крови печень забирает глюкозу и фиксирует её в виде гликогена, при снижении она «мобилизует» гликоген, переводя его через глюкозо-1-фосфат в глюкозо-6-фосфат и далее с помощью фермента фосфатазы (глюкозо-6-фосфатазы) в свободную глюкозу и фосфорную кислоту. При этом не образуется декстринов и мальтозы, что характерно для гидролитического расщепления гликогена. Одновременно в печени усиливается глюконеогенез, т.е. образование глюкозы главным образом из безазотистых остатков некоторых аминокислот. При помощи этих противоположных процессов печень поддерживает постоянство уровня сахара в крови. Процесс синтеза и распада гликогена в печени осуществляется при регулирующем воздействии гормонов на аденилат – и гуанилатциклазные механизмы гепатоцитов, с
200 | 7. Обмен углеводов |
помощью которых меняется активность внутриклеточных ферментов, принимающих участив в углеводном обмене(фосфорилаза, гликогенсинтетаза). В фиксировании глюкозы в печени на первом месте по значению стоит гексокиназа (глюкокиназа), катализирующая фосфорилирование глюкозы в глюкозо- 6-фосфат и тем самым снижающая уровень свободной глюкозы в крови. Уровень сахара в крови (норма 80-120 мг%; 3,33-5,55 ммоль/л) влияет на скорость образования глюкозо-6-фосфата путем изменения активности гексокиназы. При гипергликемии (т.е. содержании глюкозы в крови выше нормы) активность гексокиназы увеличивается; при гипогликемии (т.е. содержании глюкозы в крови ниже нормы) она, наоборот, понижается.
Таким образом, ферменты обмена углеводов выполняют регуляторные функции по поддержанию постоянства уровня глюкозы в крови, меющего важное физиологическое значение. Гипогликемия создает острую опасность нарушения снабжения центральной нервной системы глюкозой, что приводит к глубокому обмороку и смерти. Гипергликемия ведет к потере глюкозы с мочой. Гипо- и гипергликемия, особенно гипергликемия, в ряде случаев является признаком патологического состояния организма.
Помимо механизмов внутриклеточной регуляции процессы обмена углеводов у человека и животных регулируются нервной системой и гормонами.
Установлено, что важную роль в регуляции углеводного обмена занимает продолговатый мозг и гипоталамическая область. Большое значение в регуляции углеводного обмена принадлежит коре больших полушарий головного мозга. Хорошо известно, что факторы психогенного характера приводят к усилению сахарообразования в печени и гипергликемии. Гипергликемия может быть вызвана условнорефлекторным путем, что также указывает на участие коры больших полушарий в регуляции углеводного обмена. Импульсы от высших метаболических центров, расположенных в гипоталамической области, распространяются по симпатическим нервам и приводят к усилению -ин креции мозговым веществом надпочечников адреналина, стимулирующего процесс глюкозообразования из гликогена в печени и повышения сахара в крови. С другой стороны, в блуждающих нервах содержатся волокна, возбуждающие инкрецию инсулина бета-клетками островков Лангерганса поджелудочной железы, что ведет к снижению уровня сахара крови.
Таким образом, регулирующее действие нервной системы на обмен углеводов осуществляется, главным образом, через воздействие на инкрецию эндокринных желез, которые с помощью гормонов регулируют активность ферментов.
Особенно важны для регуляции углеводного обмена такие эндокринные железы, как поджелудочная железа, кора и мозговое вещество надпочечников, передняя доля гипофиза, щитовидная железа.
7. Обмен углеводов | 201 |
Инсулин – гормон β-клеток поджелудочной железы– единственный гормон, снижающий уровень сахара в крови. Его действие распространяется на мышечную и жировую ткани, а также на печень. Инсулин в печени в присутствии углеводов обладает гликогеностатическим действием. При недостаточности инсулина наблюдается повышение уровня глюкозы в крови(гипергликемия), избыточное выделение глюкозы с мочой (глюкозурия) и понижение содержания гликогена в печени. Мышечная и печеночная ткани утрачивают способность усваивать глюкозу. Одновременно подавляется биосинтез жирных кислот из глюкозы и ацетата и биосинтез белков. Недостаточность инсулина приводит к резкому снижению активности глюкокиназы и уридиндифосфотрансгликозидазы в печени. Наблюдается усиленный синтез ферментов, участвующих в глюконеогенезе из аминокислот и последний возрастает. Описанная картина недостаточности инсулина особенно сильно выражена при перерождении островков Лангерганса поджелудочной железы, в β-клетках которых вырабатывается инсулин. Это заболевание получило название диабета или сахарного мочеизнурения.
Если в печени действие инсулина на углеводный обмен развивается медленно, то в мышцах проявляется сравнительно быстро. Здесь этот гормон ускоряет метаболизм глюкозы. Он повышает окисление глюкозы и образование гликогена. Тонкий механизм действия инсулина в мышцах связывается прежде всего с повышением проницаемости клеточных мембран для глюкозы. При недостаточности инсулина замедленное поступление глюкозы в мышечную клетку, вероятно, лимитирует ее метаболизм. С повышением проницаемости мембран, вызванной инсулином, возрастает поступление глюкозы в мышечные клетки и одновременно увеличивается скорость ее окисления. Полагают, что инсулин участвует в регуляции синтеза мембранных систем переноса глюкозы и других метаболитов, увеличивая скорость образования специфических информационных РНК на рибосомах, кодирующих эти ферментные системы.
Гормоны коркового слоя надпочечников, влияющие на углеводный обмен (глюкокортикоидные гормоны: гидрокортизон, кортизон, кортикостерон), также действуют в печени и мышцах. Эти гормоны служат как бы антагонистами инсулина, их повышающее действие на уровень сахара крови основано на: увеличении скорости глюконеогенеза в печени и замедлении обмена глюкозы в периферических органах. Глюкокортикоидные гормоны повышают катаболизм белков в печени и тем самым способствуют образованию глюкозы из безазотистых фрагментов аминокислот(глюконеогенез). Под влиянием этих гормонов происходит также активирование фосфатазы(глюкозо-6-фосфа- тазы), благодаря чему из глюкозо-6-фосфата усиливается образование свободной глюкозы и ее выход из печени. На периферии, т.е. в мышцах, глюкокортикоидные гормоны замедляют окисление глюкозы и препятствуют образованию гликогена. По-видимому, это связано со способностью гормонов влиять на
Где откладываются углеводы у растений?
Рассмотрим углеводы в растениях, которые, как и жиры, органические кислоты и дубильные вещества имеют важное значение, и постоянно встречаются как в вегетативных органах, так и в органах размножения. Углеводы состоят из углерода, водорода и кислорода. Последние два элемента находятся между собой в таком же количественном сочетании, как в воде (Н2О), то есть на определенное число атомов водорода приходится в два раза меньшее число атомов кислорода. Углеводы составляют до 85-90% веществ, входящих в растительный организм. Углеводы являются основным питательным и опорным материалом в клетках и тканях растений. Углеводы подразделяются на моносахариды, дисахариды и полисахариды. Из моносахаридов в растениях распространены гексозы, имеющие состав С6Н12О6. К ним относятся глюкоза, фруктоза и др. Глюкоза (иначе называется декстроза или виноградный сахар) содержится в ягодах винограда – около 20%, в яблоках, грушах, сливах, черешне и винных ягодах. Глюкоза обладает способностью выкристаллизовываться. Фруктоза (иначе называется левулеза или плодовый сахар) кристаллизуется с трудом, встречается вместе с глюкозой в плодах, нектарниках, пчелином меде, луковицах и т. п. (Левулезой фруктоза называется потому, что при прохождении через нее поляризованного луча света последний отклоняется влево. В противоположность фруктозе виноградный сахар отклоняет поляризованный луч вправо. Поляризованным светом называется свет, пропущенный через призмы из исландского шпата, обладающего двойным лучепреломлением. Призмы эти являются составной частью поляризационного аппарата.) Свойства гексоз следующие. Они обладают особо сладким вкусом и легкорастворимы в воде. Первичное образование гексоз происходит в листьях. Они легко превращаются в крахмал, который, в свою очередь, легко может переходить в сахар при участии фермента диастаза. Глюкоза и фруктоза обладают способностью легко проникать из клетки в клетку и быстро передвигаться по растению. В присутствии дрожжей гексозы легко бродят и превращаются в спирт. Характерный и чувствительный реактив на гексозы – синяя фелингова жидкость, с помощью ее можно легко открыть малейшие их количества: при нагревании выпадает кирпично-красный осадок закиси меди. Иногда гексозы встречаются в растениях в соединении с ароматическими спиртами, с горькими или едкими веществами. Эти соединения называют тогда глюкозидами, например амигдалин, придающий горечь семенам миндаля и других косточковых растений. Амигдалин содержит ядовитое вещество – синильную кислоту. Глюкозиды не только защищают семена и плоды от поедания животными, но и предохраняют семена сочных плодов от преждевременного прорастания. Дисахариды – углеводы, имеющие состав C12h32O11. К ним относятся сахароза, или тростниковый сахар, и мальтоза. Сахароза образуется в растениях из двух частиц гексоз (глюкозы и фруктозы) с выделением частицы воды: C6h22О6 + C6h22О6 = C12h32O11 + Н2О. При кипячении с серной кислотой к тростниковому сахару присоединяется частица воды, и дисахарид распадается на глюкозу и фруктозу: C12h32О11 + Н2О = C6h22О6 + C6h22О6. Эта же реакция происходит при действии на тростниковый сахар фермента инвертазы, поэтому превращение тростникового сахара в гексозы называется инверсией, а полученные гексозы – инвертированным, сахаром. Тростниковый сахар – это тот сахар, который употребляется в пищу. Его издавна добывают из стеблей злака – сахарного тростника (Saccharum officinarum), растущего в тропических странах. Он встречается также в корнях многих корнеплодов, из которых больше всего его находится в корнях сахарной свеклы (от 17 до 23%). Из сахарной свеклы тростниковый сахар добывают на свеклосахарных заводах. Сахароза легко растворяется в воде и хорошо кристаллизуется (сахарный песок). Она не восстан
во всех частях растения, растение практически полностью состоит из углеводов, если считать по органике.
Углеводы и их роль в жизнедеятельности клетки. Видеоурок. Биология 10 Класс
Углеводы, относящиеся к органическим веществам, считаются важным элементом этого класса. Они встречаются практически везде в окружающей нас природе. На этом уроке мы узнаем, на какие классы делятся углеводы и какую роль они выполняют.
Все вещества, входящие в состав организма, делятся на два класса: органические и неорганические соединения (см. Рис. 1).
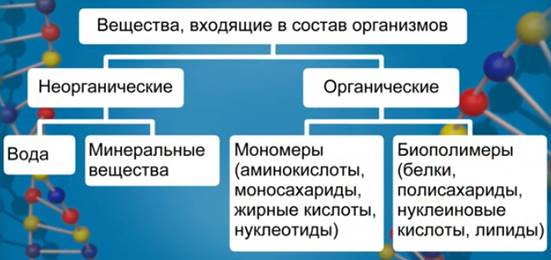
Рис. 1. Вещества, входящие в состав организмов
К неорганическим соединениям относятся вода и минеральные вещества.
К органическим соединениям относятся:
— малые органические соединения (мономеры), молекулярная масса которых колеблется от 100 до 350. Например: моносахариды, жирные кислоты, нуклеотиды. Малые органические молекулы присутствуют в клетке как в свободном виде, так и в связанном виде, то есть входят в состав биополимеров.
— большие органические молекулы (биополимеры), молекулярная масса которых колеблется от 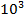 до
до 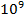 . Это белки, полисахариды, нуклеиновые кислоты, липиды.
. Это белки, полисахариды, нуклеиновые кислоты, липиды.
Углеводы – важный класс органических соединений, который встречается повсеместно: в растительных организмах, животных организмах и микроорганизмах. В растительных организмах на долю углеводов приходится 80-90 %, в животных организмах – 1-5 %, в микроорганизмах – 12-30 %.
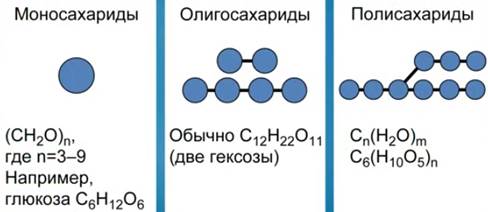
Рис. 2. Классификация углеводов
Различают три основных класса углеводов: моносахариды, олигосахариды и полисахариды (см. Рис. 2).
Бесцветные, кристаллические вещества, легко растворимые в воде и имеющие сладкий вкус.
Из моносахаридов наибольшее значение для живых организмов имеют:
1. Рибоза (см. Рис. 3). Входит в состав нуклеиновых кислот РНК, АТФ.
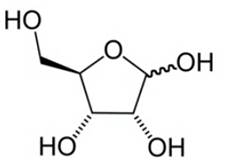
Рис. 3. Рибоза
2. Дезоксирибоза (см. Рис. 4). Входит в состав ДНК.

Рис. 4. Дезоксирибоза
3. Глюкоза (см. Рис. 5). Один из наиболее распространенных природных сахаров, находится как в свободном, так и в связанном виде. В свободном виде быстро увлекается в энергетический метаболизм, то есть служит основным источником энергии в клетке. Является мономером многих олигосахаридов и полисахаридов, например крахмала и тростникового сахара.
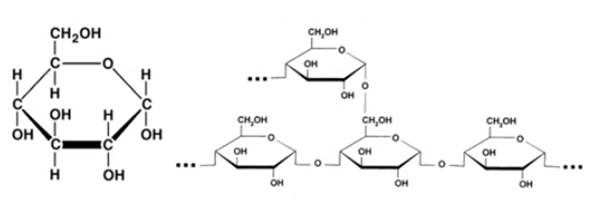
Рис. 5. Глюкоза
4. Галактоза (см. Рис. 6). Входит в состав молочного сахара – лактозы.
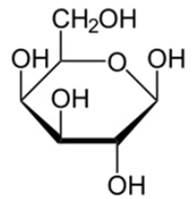
Рис. 6. Галактоза
5. Фруктоза (см. Рис. 7). Входит в состав олигосахаридов, например сахарозы. В свободном виде содержится в клетках растений.

Рис. 7. Фруктоза
Сахароподобные вещества, которые характеризуются сравнительно небольшой молекулярной массой, хорошей растворимостью в воде, легкой кристаллизацией, сладким вкусом. Количество структурных единиц, которые входят в состав олигосахаридов, – от двух до десяти.
Из олигосахаридов наиболее широко распространены дисахариды:
1. Сахароза (тростниковый сахар) – сахар, который люди употребляют в повседневной жизни (см. Рис. 8). Сахароза содержится в большинстве растений, но особенно много ее в сахарном тростнике и сахарной свекле.
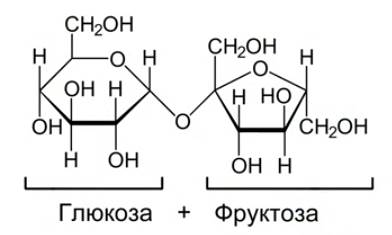
Рис. 8. Сахароза
2. Лактоза (молочный сахар) (см. Рис. 9). Содержится в молоке и молочных продуктах.
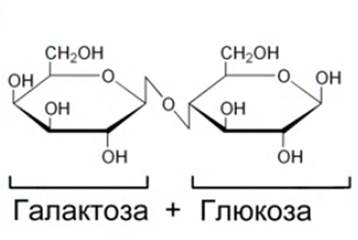
Рис. 9. Лактоза
3. Мальтоза (солодовый сахар) (см. Рис. 10). В большом количестве содержится в проросших или прорастающих зернах ячменя, ржи и пшеницы.
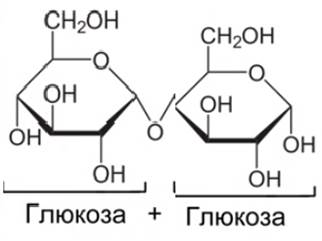
Рис. 10. Мальтоза
Являются высокомолекулярными веществами, состоящими из остатков моносахаров со степенью полимеризации выше 10. То есть количество моносахаридных звеньев может составлять до нескольких сотен или тысяч.
Из полисахаридов наибольшее значение для живых организмов имеют крахмал, гликоген, целлюлоза, хитин. Эти полисахариды не сладкие, не растворимы или плохо растворимы в воде, не кристаллизуются. Они играют роль резерва пищи и энергии (крахмал и гликоген), используются в качестве строительного материала (целлюлоза, хитин).
1. Крахмал (см. Рис. 11) – основной полисахарид в клетках растений. Он построен из остатков глюкозы. Организм человека хорошо усваивает крахмал, в составе зерновых и картофеля он потребляется в огромных количествах.
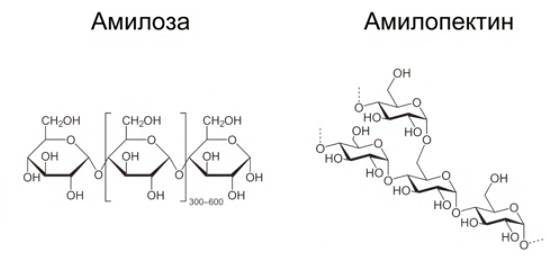
Рис. 11. Крахмал
2. Гликоген (см. Рис. 12) – полисахарид животного происхождения. Построен из остатков глюкозы. Гликоген у человека накапливается в печени и мышцах.
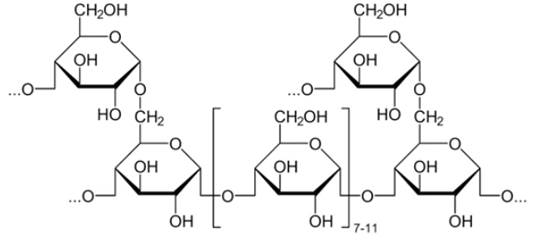
Рис. 12. Гликоген
3. Целлюлоза (см. Рис. 13) представляет собой линейный полисахарид, построенный из остатков глюкозы. Из целлюлозы построены клеточные стенки растений, и она выполняет структурную функцию.
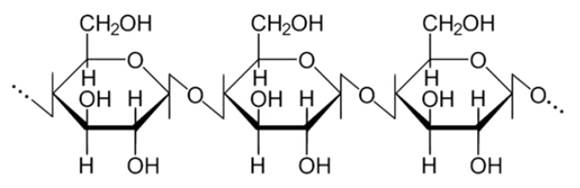
Рис. 13. Целлюлоза
4. Хитин (см. Рис. 14) – это азотосодержащий полисахарид (аминополисахарид). Хитин является вторым после целлюлозы по распространенности структурным полисахаридом. По химическому строению, физико-химическим свойствам и выполняемым функциям хитин близок к целлюлозе. Хитин – это аналог целлюлозы в животном мире.
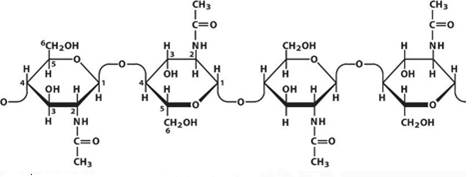
Рис. 14. Хитин (Источник)
Кленовый сироп (см. Рис. 15) – это сконцентрированный сок сахарного клена. Представляет собой раствор сахаров, в котором преобладает сахароза (65 %), а в небольших количествах содержится глюкоза и фруктоза.

Рис. 15. Кленовый сироп
Сок собирают из отверстий, проделанных в стволе дерева ранней весной. Его вытесняет диоксид углерода, образующийся в результате ряда процессов метаболизма и выделяющийся из раствора, когда дерево прогревается на весеннем солнце. Коричневый цвет кленового сиропа обуславливается не только наличием сахарозы, но и наличием аминокислот.
Многие люди, у которых по генетическим причинам отсутствует фермент лактаза, не могут усваивать молоко, так как они не могут разрушить лактозу (молочный сахар). В организме взрослого человека наличие этого фермента – скорее исключение, чем правило. Такой фермент типичен для жителей Северной Европы, в отличие от уроженцев Африки и Азии.
При попадании лактозы в неразрушенном состоянии в толстый кишечник на нее «набрасываются» бактерии, которые используют ее в пищу. В результате этого в кишечнике образовываются различные газы, происходит увеличение давления, и возникают кишечные расстройства.
Хотя целлюлоза, как и крахмал, является полисахаридом, человеческий организм ее усваивать не может, так как в организме человека отсутствует фермент целлюляза. Этот фермент перерабатывает целлюлозу.
Жвачные животные, которые постоянно используют траву в качестве пищи, способны переваривать целлюлозу с помощью различных микроорганизмов, проживающих в желудке и выделяющих целлюлязу.
Кролики, которые питаются грубой растительной пищей, изобрели способ многократного переваривания целлюлозы путем заглатывания собственных экскрементов.
1. Энергетическая
Углеводы обеспечивают до 70 % потребности организма в энергии. При окислении 1 г углеводов выделяется 17,6 кДж энергии.
2. Запасающая
Крахмал и гликоген являются запасными полисахаридами. Они являются временным хранилищем глюкозы.
3. Структурная
Целлюлоза и ряд других полисахаридов используются в качестве строительного материала. Целлюлоза входит в состав клеточных стенок растений, хитин входит в состав клеточных стенок грибов, а также используется для построения наружного скелета у членистоногих.
4. Защитная
Например, камеди (смолы, выделяющиеся при повреждении стволов и веток растений), препятствующие проникновению в раны болезнетворных микроорганизмов, являются производными моносахаридов.
Список литературы
- Каменский А.А., Криксунов Е.А., Пасечник В.В. Общая биология 10-11 класс Дрофа, 2005.
- Биология. 10 класс. Общая биология. Базовый уровень / П.В. Ижевский, О.А. Корнилова, Т.Е. Лощилина и др. – 2-е изд., переработанное. – Вентана-Граф, 2010. – 224 стр.
- Беляев Д.К. Биология 10-11 класс. Общая биология. Базовый уровень. – 11-е изд., стереотип. – М.: Просвещение, 2012. – 304 с.
- Агафонова И.Б., Захарова Е.Т., Сивоглазов В.И. Биология 10-11 класс. Общая биология. Базовый уровень. – 6-е изд., доп. – Дрофа, 2010. – 384 с.
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
- School.xvatit.com (Источник).
- Medbookaide.ru (Источник).
- Biouroki.ru (Источник).
- Sbio.info (Источник).
Домашнее задание
- Вопросы в конце параграфа 9 (стр. 37) – Каменский А.А., Криксунов Е.А., Пасечник В.В. «Общая биология», 10-11 класс
- Как подразделяются вещества, входящие в состав организмов?
Фотосинтез — Википедия
 Схематическое изображение процесса фотосинтеза, происходящего в растениях
Схематическое изображение процесса фотосинтеза, происходящего в растениях  Уравнение типа фотосинтеза, происходящего в растениях
Уравнение типа фотосинтеза, происходящего в растенияхФотоси́нтез (от др.-греч. φῶς — свет и σύνθεσις — соединение, складывание, связывание, синтез) — сложный химический процесс преобразования энергии света (в некоторых случаях инфракрасного излучения) в энергию химических связей органических веществ при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл у бактерий и бактериородопсин у архей). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндергонических реакциях, в том числе превращения углекислого газа в органические вещества.
У живых организмов обнаружено два типа пигментов, способных выполнять функцию фотосинтетических антенн. Эти пигменты поглощают кванты видимого света и обеспечивают дальнейшее запасание энергии излучения в виде энергии электрохимического градиента H+ на биологических мембранах. У подавляющего большинства организмов роль антенн играют хлорофиллы; менее распространён случай, при котором в качестве антенны служит производное витамина А ретиналь. В соответствии с этим выделяют хлорофилльный и бесхлорофилльный фотосинтез.
Бесхлорофилльный фотосинтез[править | править код]
Система бесхлорофилльного фотосинтеза отличается значительной простотой организации, в связи с чем предполагается эволюционно первичным механизмом запасания энергии электромагнитного излучения. Эффективность бесхлорофилльного фотосинтеза как механизма преобразования энергии сравнительно низка (на один поглощённый квант переносится лишь один H+).
Открытие у галофильных архей[править | править код]
Dieter Oesterhelt и Walther Stoeckenius идентифицировали в «пурпурных мембранах» представителя галофильных архей Halobacterium salinarium (прежнее название Н. halobium) белок, который позже был назван бактериородопсином. Впоследствии были накоплены факты, указывающие на то, что бактериородопсин является светозависимым генератором протонного градиента. В частности, было продемонстрировано фотофосфорилирование на искусственных везикулах, содержащих бактериородопсин и митохондриальную АТФ-синтазу, фотофосфорилирование в интактных клетках H. salinarium, светоиндуцируемое падение pH среды и подавление дыхания, причём все эти эффекты коррелировали со спектром поглощения бактериородопсина. Таким образом, были получены неопровержимые доказательства существования бесхлорофилльного фотосинтеза.
Механизм[править | править код]
Механизм бесхлорофилльного фотосинтеза галобактерийФотосинтетический аппарат экстремальных галобактерий является наиболее примитивным из ныне известных; в нём отсутствует электронтранспортная цепь. Цитоплазматическая мембрана галобактерий является сопрягающей мембраной, содержащей два основных компонента: светозависимый протонный насос (бактериородопсин) и АТФ-синтазу. Работа такого фотосинтетического аппарата основана на следующих трансформациях энергии:
- Хромофор бактериородопсина ретиналь поглощает кванты света, что приводит к конформационным изменениям в структуре бактериородопсина и транспорту протона из цитоплазмы в периплазматическое пространство. Таким образом, в результате работы бактериородопсина энергия солнечного излучения трансформируется в энергию электрохимического градиента протонов на мембране.
- При работе АТФ-синтазы энергия трансмембранного градиента трансформируется в энергию химических связей АТФ. Таким образом осуществляется хемиосмотическое сопряжение.
При бесхлорофилльном типе фотосинтеза (как и при реализации циклических потоков в электрон-транспортных цепях) не происходит образования восстановительных эквивалентов (восстановленного ферредоксина или НАД(Ф)Н), необходимых для ассимиляции углекислого газа. Поэтому при бесхлорофилльном фотосинтезе не происходит ассимиляции углекислого газа, а осуществляется исключительно запасание солнечной энергии в форме АТФ (фотофосфорилирование).
Значение[править | править код]
Основной путь получения энергии для галобактерий — аэробное окисление органических соединений (при культивировании используют углеводы и аминокислоты). При дефиците кислорода помимо бесхлорофилльного фотосинтеза источниками энергии для галобактерий может служить анаэробное нитратное дыхание или сбраживание аргинина и цитруллина. Однако в эксперименте было показано, что бесхлорофилльный фотосинтез может служить и единственным источником энергии в анаэробных условиях при подавлении анаэробного дыхания и брожения при обязательном условии, что в среду вносят ретиналь, для синтеза которого необходим кислород.
Хлорофилльный фотосинтез[править | править код]
Хлорофилльный фотосинтез отличается от бактериородопсинового значительно большей эффективностью запасания энергии. На каждый поглощённый квант излучения против градиента переносится не менее одного H+, и в некоторых случаях энергия запасается в форме восстановленных соединений (ферредоксин, НАДФ).
Аноксигенный[править | править код]
Аноксигенный (или бескислородный) фотосинтез протекает без выделения кислорода. К аноксигенному фотосинтезу способны пурпурные и зелёные бактерии, а также гелиобактерии.
При аноксигенном фотосинтезе возможно осуществление:
- Светозависимого циклического транспорта электронов, не сопровождающегося образованием восстановительных эквивалентов и приводящего исключительно к запасанию энергии света в форме АТФ. При циклическом светозависимом электронном транспорте необходимости в экзогенных донорах электронов не возникает. Потребность в восстановительных эквивалентах обеспечивается нефотохимическим путём, как правило, за счёт экзогенных органических соединений.
- Светозависимого нециклического транспорта электронов, сопровождающегося и образованием восстановительных эквивалентов, и синтезом АДФ. При этом возникает потребность в экзогенных донорах электронов, которые необходимы для заполнения электронной вакансии в реакционном центре. В качестве экзогенных доноров электронов могут использоваться как органические, так и неорганические восстановители. Среди неорганических соединений наиболее часто используются различные восстановленные формы серы (сероводород, молекулярная сера, сульфиты, тиосульфаты, тетратионаты, тиогликоляты), также возможно использование молекулярного водорода.
Оксигенный[править | править код]
Оксигенный (или кислородный) фотосинтез сопровождается выделением кислорода в качестве побочного продукта. При оксигенном фотосинтезе осуществляется нециклический электронный транспорт, хотя при определённых физиологических условиях осуществляется исключительно циклический электронный транспорт. В качестве донора электронов при нециклическом потоке используется крайне слабый донор электронов — вода.
Оксигенный фотосинтез распространён гораздо шире. Характерен для высших растений, водорослей, многих протистов и цианобактерий.
Фотосинтез — процесс с крайне сложной пространственно-временной организацией.
Разброс характерных времен различных этапов фотосинтеза составляет 19 порядков: скорость процессов поглощения квантов света и миграции энергии измеряется в фемтосекундном интервале (10−15 с), скорость электронного транспорта имеет характерные времена 10−10—10−2 с, а процессы, связанные с ростом растений, измеряются днями (105—107 с).
Также большой разброс размеров характерен для структур, обеспечивающих протекание фотосинтеза: от молекулярного уровня (10−27 м3) до уровня фитоценозов (105 м3).
В фотосинтезе можно выделить отдельные этапы, различающиеся по природе и характерным скоростям процессов:
- фотофизический;
- фотохимический;
- химический:
- реакции транспорта электронов;
- «темновые» реакции или циклы углерода при фотосинтезе.
На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы (пластохинон).[1].
На втором этапе происходит разделение зарядов в реакционном центре. Молекула воды теряет электрон под воздействием катиона-радикала, образовавшегося из молекулы хлорофилла после потери ей своего электрона и передачи его пластохинону на первом этапе: h3O − e−⟶H++O˙H{\displaystyle {\ce {h3O\ -\ e^{-}->H^{+}{}+{\dot {O}}H}}}. Затем образовавшиеся гидроксильные радикалы под воздействием положительно заряженных ионов марганца преобразуются в кислород и воду: 4O˙H⟶O2+2h3O{\displaystyle {\ce {4{\dot {O}}H->O2{}+2h3O}}}[1]. Одновременно с этим процессом происходит перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН[1]. Первые два этапа вместе называют светозависимой стадией фотосинтеза.
Третий этап заключается в поглощении второй молекулой хлорофилла кванта света и передаче ею электрона ферредоксину. Затем хлорофилл получает электрон после цепи его перемещений на первом и втором этапах. Ферредоксин восстанавливает универсальный восстановитель НАДФ.[1].
Четвёртый этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха[1].
Лист[править | править код]
Фотосинтез растений осуществляется в хлоропластах — полуавтономных двухмембранных органеллах, относящихся к классу пластид. Хлоропласты могут содержаться в клетках стеблей, плодов, чашелистиков, однако основным органом фотосинтеза является лист. Он анатомически приспособлен к поглощению энергии света и ассимиляции углекислоты. Плоская форма листа, обеспечивающая большое отношение поверхности к объёму, позволяет более полно использовать энергию солнечного света. Вода, необходимая для поддержания тургора и протекания фотосинтеза, доставляется к листьям из корневой системы по ксилеме — одной из проводящих тканей растения. Потеря воды в результате испарения через устьица и в меньшей степени через кутикулу (транспирация) служит движущей силой транспорта по сосудам. Однако избыточная транспирация является нежелательной, и у растений в ходе эволюции сформировались различные приспособления, направленные на снижение потерь воды. Отток ассимилятов, необходимый для функционирования цикла Кальвина, осуществляется по флоэме. При интенсивном фотосинтезе углеводы могут полимеризоваться, и при этом в хлоропластах формируются крахмальные зёрна. Газообмен (поступление углекислого газа и выделение кислорода) осуществляется путём диффузии через устьица (некоторая часть газов движется через кутикулу).
Поскольку дефицит углекислого газа значительно увеличивает потери ассимилятов при фотодыхании, необходимо поддерживать высокую концентрацию углекислоты в межклеточном пространстве, что возможно при открытых устьицах. Однако поддержание устьиц в открытом состоянии при высокой температуре приводит к усилению испарения воды, что приводит к водному дефициту и также снижает продуктивность фотосинтеза. Этот конфликт решается в соответствии с принципом адаптивного компромисса. Кроме того, первичное поглощения углекислого газа ночью, при низкой температуре, у растений с CAM-фотосинтезом позволяет избежать высоких транспирационных потерь воды.
У некоторых суккулентов с вырожденными листьями (например, у большинства видов кактусов) основная фотосинтетическая активность связана со стеблем.
Фотосинтез на тканевом уровне[править | править код]
На тканевом уровне фотосинтез у высших растений обеспечивается специализированной тканью — хлоренхимой. Она располагается близ поверхности тела растения, где получает достаточно световой энергии. Обычно хлоренхима находится непосредственно под эпидермой. У растений, растущих в условиях повышенной инсоляции, между эпидермой и хлоренхимой может располагаться один или два слоя прозрачных клеток (гиподерма), обеспечивающих рассеивание света. У некоторых тенелюбивых растений хлоропластами богата и эпидерма (например, кислица). Часто хлоренхима мезофилла листа дифференцирована на палисадную (столбчатую) и губчатую, но может состоять и из однородных клеток. В случае дифференцировки наиболее богата хлоропластами палисадная хлоренхима.
 Хлоропласты в клетках листа
Хлоропласты в клетках листаХлоропласты[править | править код]
Хлоропласты отделены от цитоплазмы двойной мембраной, обладающей избирательной проницаемостью. Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые, соединяясь друг с другом, образуют тилакоиды, которые, в свою очередь, группируются в стопки, называемые гранами. На мембранах тилакоидов располагаются молекулы хлорофилла и других вспомогательных пигментов (каротиноиды). Поэтому их называют фотосинтезирующими мембранами. Внутритилакоидное пространство отделено и не сообщается с остальной стромой; предполагается также, что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (типа 70s), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Фотосинтетические мембраны прокариот[править | править код]
Цианобактерии, таким образом, как бы сами являются хлоропластом, и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФ, использующийся как восстановитель. В качестве побочного продукта выделяется кислород.
Фотохимическая суть процесса[править | править код]
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённые состояния, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно, и хлорофилл за 10−12 с переходит с него на первое с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция и фосфоресценция соответственно) или тепла с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где под воздействием кванта света переходящий в возбуждённое состояние хлорофилл становится донором электрона (восстановителем) и передаёт его первичным акцепторам. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше, чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделение зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным же донором в случае оксигенного фотосинтеза является вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции H2O → O2 (E0 = +0,82 В) и НАДФ+ (E0 = −0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больше +0,82 В, чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньше −0,32 В, чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировались две фотосистемы, и для полного проведения процесса необходимы два кванта света и два хлорофилла разных типов.
Светособирающие комплексы[править | править код]
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла, в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каротиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
Основные компоненты цепи переноса электронов[править | править код]
Фотосистема II[править | править код]
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла-a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется, и возбуждённая молекула П680 становится сильным восстановителем (химический потенциал E0 = −0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к комплексу b6f. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства, и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
Цитохром-b6/f-комплекс[править | править код]
b6f-комплекс — это насос, перекачивающий протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Фотосистема I[править | править код]
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f-комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона[править | править код]
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f-комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
В темновой стадии с участием АТФ и НАДФ происходит восстановление CO2 до глюкозы (C6H12O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3-фотосинтез, цикл Кальвина[править | править код]
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- карбоксилирования;
- восстановления;
- регенерация акцептора CO2.
На первой стадии к рибулозо-1,5-бисфосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа. Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы с образованием 1,3-дифосфоглицериновой кислоты (ДФГК), затем при воздействии триозофосфатдегидрогеназы и НАДФН ацил-фосфатная группа ДФГК дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат — фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4-фотосинтез[править | править код]
Отличие этого механизма фотосинтеза от обычного заключается в том, что фиксация углекислого газа и его использование разделены в пространстве между различными клетками растения[2].
При низкой концентрации растворённого в строме CO2рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения типа С4 изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4-фотосинтез практически не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
Фотосинтез по пути С4 проводят около 7600 видов растений. Все они относятся к цветковым: многие Злаковые (61 % видов, в том числе культурные — кукуруза, сахарный тростник и сорго и др.[3][4]), Гвоздичноцветные (наибольшая доля в семействах Маревые — 40 % видов, Амарантовые — 25 %), некоторые Осоковые, Астровые, Капустные, Молочайные[5][6].
CAM-фотосинтез[править | править код]
При фотосинтезе типа CAM (англ. Crassulaceae acid metabolism — кислотный метаболизм толстянковых) происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве, как у С4, а во времени[2]. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.

Фотосинтез составляет энергетическую основу всего живого на планете, кроме хемосинтезирующих бактерий.
Фотосинтез совершается в зеленых частях наземных растений и в водорослях. За один год зеленые водоросли выделяют в атмосферу Земли 3,6∗1011{\displaystyle 3,6*10^{11}} тонн кислорода, что составляет 90%{\displaystyle 90\%} всего кислорода, вырабатываемого в процессе фотосинтеза на Земле. Фотосинтез — самый крупный биохимический процесс на Земле.[7]
Возникновение на Земле более 3 млрд лет назад механизма расщепления молекулы воды квантами солнечного света с образованием O2 представляет собой важнейшее событие в биологической эволюции, сделавшее свет Солнца главным источником энергии биосферы.
Фототрофы обеспечивают конверсию и запасание энергии термоядерных реакций, протекающих на Солнце, в энергию органических молекул. Солнечная энергия при участии фототрофов конвертируется в энергию химических связей органических веществ. Существование гетеротрофных организмов возможно исключительно за счёт энергии, запасённой фототрофами в органических соединениях. При использовании энергии химических связей органических веществ гетеротрофы высвобождают её в процессах дыхания и брожения.
Фотосинтез является основой продуктивности как сельскохозяйственных растений, так и животной пищи.
Энергия, получаемая человечеством при сжигании биотоплива (дрова, пеллеты, биогаз, биодизель, этанол, метанол) и ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез служит главным входом неорганического углерода в биогеохимический цикл.
Большая часть свободного кислорода атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни существовать на суше.
Первые опыты по изучению фотосинтеза были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал поддерживать горение, а помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод, что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил, что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено, что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 году Роберт Майер на основании закона сохранения энергии постулировал, что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 году В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы были впервые выделены в 1818 году П. Ж. Пеллетье и Ж. Каванту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал, что именно поглощённые лучи позволяют повысить энергию системы, создав вместо слабых связей С−О и О−Н высокоэнергетические С−С (до этого считалось, что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2: в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось, что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Большой вклад в изучение фотосинтеза внёс русский учёный Андрей Сергеевич Фаминцын.[8] В 1868 году он впервые экспериментально доказал и научно обосновал применение искусственного освещения для выращивания растений, использовав керосиновые лампы вместо солнечного света.[9] Фамицын первым системно изучил процессы образования крахмала в тканях растений под воздействием света, а также влияния света на образование хлорофилла, его расположение в листьях растений различных таксонов.[10][11]
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль, он же в 1931 году доказал, что пурпурные бактерии и зелёные серобактерии осуществляют аноксигенный фотосинтез[12][13]. Окислительно-восстановительный характер фотосинтеза означал, что кислород в оксигенном фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 году А. П. Виноградов в опытах с изотопной меткой. В 1937 году Роберт Хилл установил, что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954—1958 годах Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мелвином Кальвином с использованием изотопов углерода в конце 1940-х годов, за эту работу в 1961 году ему была присуждена Нобелевская премия.
В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа. С4-фотосинтез был описан Ю. С. Карпиловым в 1960 году и М. Д. Хэтчем и Ч. Р. Слэком в 1966 году.
Существующие данные указывают на то, что наиболее древними организмами, запасающими энергию света в форме химической энергии, были археи, осуществляющие бесхлорофилльный фотосинтез, при котором не происходит образования восстановителя (НАДФН) и фиксации углекислого газа, а энергия запасается лишь в форме АТФ.
Несколько позже (3,7—3,8 миллиарда лет назад) независимо от архей и многократно в ходе эволюции возникали организмы (зелёные, пурпурные бактерии и другие группы эубактерий) с одной из фотосистем, позволяющей осуществлять аноксигенный фотосинтез, при котором в качестве доноров электронов используются соединения с низким редокс-потенциалом (водород, сульфиды и сероводород, сера, соединения железа (II), нитриты) или осуществляется циклический поток электронов вокруг фотосистемы.
Система оксигенного фотосинтеза, при которой в электрон-транспортной цепи совместно функционируют две фотосистемы, характерная для цианобактерий и происшедших от них в ходе эндосимбиоза хлоропластов водорослей и высших растений, возникла в эволюции однократно, по разным оценка
Образование углеводов в процессах фотосинтеза и хемосинтеза — Мегаобучалка
Фотосинтез. Фотосинтез — это совокупность процессов, в ходе которых запасается солнечная энергия в виде химических связей органических соединений, синтезируемых из неорганических веществ.
Рис. 15. Строение хлорофилла а: I—IV — пиррольные циклы; пунктиром показаны координационные связи
Фотосинтез состоит из двух фаз: световой (фотофизический и фотохимический этапы) и темновой. В ходе световой фазы происходит поглощение солнечной энергии хлорофиллом и передача ее в «реакционный центр», где в результате химических реакций, включающих транспорт электронов между различными переносчиками и сопряженного с ним фосфорилирования, образуются восстановительные и энергетические эквиваленты (НАДФН и АТФ). Для нормального функционирования реакций световой фазы кроме света необходимы хлорофилл и вода (или другой источник водорода). Различные хлорофиллы способны поглощать кванты света с определенной длиной волны и переходить при этом в возбужденное состояние. Переход в исходное состояние приводит к высвобождению энергии, которая через ряд промежуточных стадий запасается в форме АТФ и НАДФН.
Все известные в настоящее время фотосинтезирующие организмы содержат хлорофиллы — зеленые магнийпорфириновые пигменты. Известно свыше десяти их видов, различающихся природой химических группу присоединенных к пиррольным структурам порфиринового ядра, окраской, распространением среди живых организмов. Так, у всех зеленых растений содержатся хлорофиллы а и b (рис 17), в диатомовых водорослях — хлорофилл с, в красных водорослях — хлорофилл d. В клетках пурпурных бактерий обнаружены бактериохлорофиллы a и b, а в зеленых бактериях — бактериохлорофиллы с и d. Важным свойством молекул хлорофилла является их способность взаимодействовать с белками и друг с другом, образуя агрегированные формы с различными спектрами поглощения.
Наряду с зелеными пигментами в хлоропластах и хроматофорах содержатся каротиноиды — желтые и оранжевые пигменты
Структура β-каротина
изопреновой природы. Каротиноиды могут быть разделены на ряд групп по своему строению: собственно каротиноиды, гидроксилсодержащие каротиноиды и каротиноиды, содержащие карбонильные группы. Основными представителями у высших растений являются β-каротин и ксантофилл. Предполагают, что каротиноиды, используя лучи, не поглощаемые хлорофиллом, передают их энергию на молекулы хлорофилла. Существуют данные, что каротиноиды предохраняют молекулы хлорофилла от разрушения в процессе фотоокисления, а также играют определенную роль в половом процессе при прорастании пыльцы и росте пыльцевых трубок, у высших растений и при созревании половой клетки у водорослей и грибов.
Третья группа пигментов — фикобилины. Это красные и синие пигменты (фикоэритрины, фикоцианины), содержащиеся в хроматофорах некоторых водорослей. В основе химического строения фикобилинов лежит та же тетрапиррольная структура, но пиррольные группы расположены линейно. Фикобилины поглощают энергию света в зеленой и желтой областях спектра и передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей — пример приспособления в ходе эволюции к поглощению тех лучей солнечного света, которые проникают через толщу морской воды.
Темновая фаза фотосинтеза — это фиксация и восстановление СО2 с образованием углеводов и других конечных продуктов фотосинтеза. На этой стадии свет не нужен, а используются образованные в световой фазе восстановительные и энергетические эквиваленты. Во время темновой фазы атомы водорода, поставляемые световыми реакциями, используются для восстановления СО2 до углеводов согласно общему уравнению фотосинтеза:
Свет
6С02 + 12Н20 → С6Н12О6 + 602 + 6Н20
Хлорофилл
При этом на каждый моль синтезированного углевода запасается ~160 кДж энергии.
Процесс восстановления СО2 начинается с его присоединения к пятиуглеродному акцептору рибулозо-1,5-бисфосфату (РуБФ). Образующееся шестиуглеродное соединение очень нестойко. Опытами М. Кальвина с меченым оксидом углерода (IV) установлено, что при длительности фотосинтеза 2 с первым фиксируемым меченым соединением является не С6-соединение, а 3-фосфоглицериновая кислота (ФГК), причем метка обнаруживается только в карбоксиле. Реакция карбоксилирования катализируется рибулозобисфосфат-карбоксилазой (РБФ-карбоксилаза). Затем образующаяся ФГК фосфорилируется при участии фермента фосфоглице-раткиназы с использованием АТФ и превращается в 1,3-бисфосфо-D-глицериновую кислоту, которая более реакционноспособна и легче восстанавливается в глицеральдегид-3-фосфат (ГАФ).
В реакции восстановления принимают участие образовавшийся в световой стадии НАДФН (никотинамидадениннуклеотид) и фермент глицеральдегидфосфатдегидрогеназа. Часть образовавшихся молекул ГАФ под действием фермента триозофосфатизомеразы превращается в дигидроксиацетонфосфат (ДГАФ). Эти два триозофосфата конденсируются под действием альдолазы. Сначала образуется фруктозо-1,6-дифосфат (ФДФ), далее — монофосфаты фруктозы (Ф6Ф) и глюкозы (Г6Ф) и, наконец, — сахароза и крахмал. Для того чтобы процесс фотосинтеза продолжался, необходима постоянная регенерация РуБФ, поэтому в одном обороте цикла участвуют 6 молекул РуБФ, которые фиксируют 6 молекул С02. Образующиеся 12 молекул ФГК, а после ее восстановления 12 молекул ГАФ расходуются следующим образом:
5ГАФ → 5ДГАФ
ЗГАФ + ЗДГАФ -+ ЗФДФ →ЗФ6Ф
2ГАФ + 2Ф6Ф-→2 Ксилулоза-5Ф + 2Эритрозо-4Ф
2ГАФ + 2 Седогептулозо-7Ф 2 Ксилулозо-5Ф + 2 Рибозо-5Ф
Таким образом, две молекулы глицеральдегид-3-фосфата образуют одну молекулу гексозы, которая выходит из цикла, а 10 идут на регенерацию шести молекул рибулозо-1,5 бисфосфата, и цикл замыкается.
У некоторых растений в качестве первичных продуктов фотосинтеза сначала образуются оксалоацетат и малат в результате карбоксилирования ФЕП под действием ФЕП-карбоксилазы. Поскольку первичные продукты в этом случае содержат четыре атома углерода, его называют С4-путь фотосинтеза в отличие от цикла Кальвина, где образующаяся ФГК содержит три атома углерода (С3-путь). С4-Путь фотосинтеза, в котором принимают участие два типа клеток и два типа хлоропластов, называется кооперативным.
Хемосинтез. Гетеротрофная фиксация С02. Известный русский микробиолог С. Н. Виноградский показал (1856—1953), что органические вещества синтезируются в природе не только путем фотосинтеза в зеленых растениях, но и бактериями, не содержащими хлорофилла. Энергию, необходимую для синтеза органических соединений, эти бактерии получают при окислении различных неорганических соединений: Fe, N, S, Н, Sb, Мn. Этот процесс называется хемосинтезом. Некоторые хемосинтетики используют в качестве доноров водорода простейшие органические вещества — метан, метанол и пр.
Исследование химизма ассимиляции меченого оксида углерода (IV) (14С02) различными хемосинтезирующими бактериями показало, что первым стойким продуктом хемосинтеза является фосфоглицериновая кислота, а присоединение СO2 к рибулозобисфосфату, т. е. цикл Кальвина, — основным механизмом ассимиляции СО2. У многих хемосинтезирующих бактерий цикл Кальвина — главный, но не единственный путь образования органических веществ.
Таким образом, процессы фотосинтеза и хемосинтеза — источники органического вещества на Земле.
А. Ф. Лебедевым еще в 1914 г. была высказана мысль, что гетеротрофные организмы могут частично ассимилировать углерод не только из готовых органических соединений, но и из СО2, связывая последний с некоторыми кетокислотами. Впоследствии это получило экспериментальное подтверждение в исследованиях А. Л. Курсанова, Г. Вуда, С. Очоа и др.
Однако, как уже указывалось, между автотрофами и гетеротрофами существует коренное отличие: первые способны синтезировать органическое вещество полностью за счет неорганических веществ (СО2 и Н20), вторые усваивают СО2, используя готовое органическое соединение, например пируват.
6.6. Превращение углеводов в процессе пищеварения
Человек и животные не способны к первичному биосинтезу углеводов из неорганических веществ, они могут лишь образовывать их в процессе глюконеогенеза из других органических веществ (органических кислот, жиров, аминокислот), но главным источником углеводов является пища. Некоторые насекомые получают с пищей преимущественно одни углеводы (питающиеся древесиной, цветочным нектаром). Они настолько приспособились к подобному питанию, что погибают без углеводов. У ряда беспозвоночных животных существует хорошо выраженная сезонная периодичность содержания углеводов в организме. Так, у мидий и креветок осенью резко увеличивается количество углеводов, происходит запасание на зиму питательных веществ. Этому способствует и повышенное содержание углеводов в планктоне — пище мидий и креветок. Углеводы составляют существенную часть пищевого рациона человека и многих животных. На их долю приходится 60—70% общей суммы калорий пищи человека. Особенно много углеводов содержат крупы и макароны (65—75%), хлеб (около 50%), картофель (до 25%). Очень мало их в мясомолочных продуктах (0,5-2%).
Углеводы всасываются через слизистую оболочку кишечника только в виде моносахаридов. Даже хорошо растворимые дисахариды сахароза и лактоза не могут в неизмененном виде всасываться в кишечнике. Если их ввести в кровь, минуя желудочно-кишечный тракт, дисахариды не будут утилизироваться клетками различных тканей. Это еще в большей степени относится к нерастворимым в воде полисахаридам: крахмалу, гликогену. Таким образом, в процессе переваривания углеводов пищи должно происходить их расщепление до моносахаридов — той единственной формы, которая может всасываться и использоваться в организме.
Переваривание углеводов начинается в ротовой полости. В составе слюны содержатся два фермента: α-амилаза и мальтаза.
Специфической особенностью α-амилазы слюны (старое название — птиалин) является способность гидролитически расщеплять крахмал только тех пищевых продуктов, которые подвергались термической обработке при изготовлении пищи. «Сырой» крахмал во рту почти не гидролизуется, только при очень медленном жевании. α-Амилаза слюны млекопитающих в отличие от птиц высоко активна. Интересно, что у некоторых приматов (бабуины, резусы) этот фермент отсутствует, хотя у человека он очень активен.
Пища в ротовой полости находится недолго, после попадания ее в желудок постепенно пищевой комок пропитывается кислым желудочным соком. Низкие значения рН инактивируют α-амилазу слюны. Таким образом, ее гидролитическое действие осуществляется кратковременно, в ротовой полости расщепление крахмала и гликогена только начинается. В желудке амилолитические ферменты отсутствуют.
Основным местом переваривания крахмала и гликогена является тонкий кишечник, где на них действует α-амилаза поджелудочной железы. Она может расщеплять крахмал, не подвергшийся термической обработке при приготовлении пищи. Поэтому появление в кале непереваренных гранул крахмала — признак либо нарушения выделения, либо нарушения функции панкреатической α-амилазы. Установлено, что активность α-амилазы в кишечнике отдельных видов рыб зависит от типа питания: у видов, питающихся фитопланктоном, который богат крахмалом, она значительно выше, чем у потребляющих зоопланктон.
В поджелудочном соке содержится также фермент мальтаза, расщепляющий дисахарид мальтозу. Однако основная масса дисахаридов, образовавшихся в результате действия α-амилазы и поступивших с пищей, гидролизуется ферментами тонкого кишечника. Этот процесс происходит не в просвете кишечника, а в клетках слизистой оболочки. Там действуют мальтаза, изомальтаза и сахараза (инвертаза). Мальтаза и изомальтаза находятся обычно в прочном комплексе с инвертазой. Эти ферменты функционируют в щеточной кайме эпителия слизистой оболочки кишечника в количествах, обеспечивающих переваривание и усвоение пищи взрослого человека. Помимо перечисленных ферментов кишечный эпителий содержит β- галактозидазу с оптимумом рН 4,5, гетерогалактозидазу и лактазу.
В результате последовательного воздействия перечисленных ферментов углеводы превращаются в моносахариды. Они хорошо всасываются кишечной стенкой. Скорость их всасывания различна: наибольшая для галактозы, близка для глюкозы, примерно половину последней составляет для фруктозы, у маннозы и ксилозы около четверти от скорости всасывания глюкозы, самая малая у арабинозы. Пентозы и манноза проникают через эпителий путем облегченной диффузии, их поступление против градиента концентрации невозможно. Интенсивность всасывания пентоз и маннозы зависит поэтому от скорости удаления этих сахаров путем диффузии наружу, на серозную сторону клеток, откуда они быстро уносятся с током крови.
У человека и млекопитающих животных отсутствует фермент целлюлаза, вызывающий гидролиз клетчатки (целлюлозы). Не атакуются ферментами желудочно-кишечного тракта млекопитающих и растительные пентозаны. Только некоторые из них частично расщепляются бактериями в толстом кишечнике с образованием органических кислот, С02 и других веществ. Поскольку многие млекопитающие растительноядны, переваривание целлюлозы для них крайне необходимо. В связи с этим они имеют специализированный пищеварительный тракт, в высокой степени приспособленный для симбиотического переваривания целлюлозы. Так, у жвачных желудок состоит из нескольких отделов, первый и самый большой из которых называется рубцом. Он содержит в больших количествах бактерии и простейших типа инфузорий, продуцирующих фермент целлюлазу. Рубец можно сравнить с бродильным чаном, в котором пища, смешанная со слюной, подвергается интенсивной ферментации. Продукты брожения (главным образом, масляная, уксусная и пропионовая кислоты), всасываются и используются организмом, С02 и СН4 выводятся с отрыжкой. Очень важно, что образовавшиеся из целлюлозы органические кислоты могут непосредственно использоваться на биосинтез жиров, они являются у жвачных главным источником энергии (у крупного рогатого скота составляют до 70% всей необходимой организму энергии).
Реакция содержимого рубца у здорового животного при нормальной организации кормления постоянно поддерживается в пределах рН 6,5-7,4 и смещается в кислую сторону в период наиболее интенсивного сбраживания корма. В этот момент образование органических кислот брожения превалирует над их всасыванием и нейтрализацией.
Среда рубца чрезвычайно благоприятна для размножения микроорганизмов. рН находится в пределах 6,5-7,4, температура колеблется от 39 до 40 °С. Кислород, который токсичен для многих видов бактерий, в рубце почти отсутствует.
Интенсивность бродильных процессов в рубце очень велика. За сутки в нем образуется до 4 л летучих жирных кислот (ЛЖК). Общее количество ЛЖК и соотношение отдельных кислот зависит от структуры рациона. В большинстве случаев в рубце преобладает уксусная кислота. Наибольшее ее количество образуется при даче рационов, богатых клетчаткой. Обилие в рационе крахмалистых и сахаристых кормов благоприятствует образованию пропионовой кислоты.
В процессе жизнедеятельности микроорганизмов в рубце образуются также газы. Они необходимы для дальнейших реакций, в результате которых синтезируется ряд ценных питательных веществ. Количество и состав газов, образующихся через 2 — Зч после кормления, достигает у крупного рогатого скота до 25-35 л/ч. За сутки может образовываться до 700 л газов.
В рубце образуются аммиак, метан, двуокись углерода, азот, небольшое количество водорода, сероводорода и кислорода.
В итоге деятельности микрофлоры рубца каждый день в нем вырабатывается до 2,5кг бактериального протеина (400 г азота). Этот бактериальный протеин переваривается в тонком кишечнике и служит главным источником аминокислот для коровы.
Улучшение переваривания растительного материала с помощью симбиотического микробного брожения встречается и у некоторых птиц, например у белой куропатки, которая несколько зимних месяцев питается в основном почками и молодыми ветками. У большинства птиц семейства куриных имеются две слепые кишки для сбраживания целлюлозы. Локализация этого процесса в заднем отделе кишечника менее выгодна, чем в рубце, так как исключается возможность дальнейшего переваривания пищи при прохождении кишечника. У многих грызунов, кроликов и зайцев это компенсируется капрофагией, поеданием кала.
6.7. Регуляция постоянства содержания глюкозы в крови
Всосавшиеся из кишечника моносахариды (преимущественно глюкоза) через воротную вену доставляются прежде всего в печень, где часть глюкозы используется на образование запасных отложений гликогена. Содержание последнего в печени человека и животных зависит от режима питания; чем больше углеводов в рационе, тем больше гликогена в печени. Синтез и распад гликогена катализируются различными ферментами и контролируются независимо. Часть глюкозы используется самой печенью для получения энергии, необходимой для многочисленных реакций, протекающих в ней. Кроме того, определенное количество глюкозы в печени превращается в жиры. Так, при нормальном смешанном питании только 3—5% глюкозы превращается в гликоген (его среднее содержание в печени колеблется от 5 до 7%), 30% глюкозы идет на образование жиров и 60—70% окисляется до С02 и Н20. При обильном углеводном питании 10% глюкозы откладывается в печени в виде гликогена, 40%—превращается в жиры и 50% окисляется.
Сахар в крови, лимфе, спинномозговой жидкости представлен глюкозой, уровень которой очень тонко регулируется. Содержание глюкозы в артериальной и капиллярной крови здорового человека составляет 54—96 мг/100 мл при определении глюкозооксидазным методом и 80—120 мг/100 мл при определении по Хагедорну и Иенсену (вторым методом кроме глюкозы определяются одновременно некоторые другие редуцирующие вещества в крови). В венозной крови содержится меньше глюкозы, так как она потребляется тканями из крови. Единственный орган, выделяющий глюкозу в общую циркуляцию, — печень.
Инсулин секретируется β-клетками островков Лангерганса поджелудочной железы в форме проинсулина. Основным проявлением недостаточности инсулина является повышение уровня глюкозы в крови (гипергликемия), избыточное выведение глюкозы с мочой (глюкозурия) и понижение содержания гликогена в печени. Недостаточность инсулина подавляет биосинтез жирных кислот из глюкозы и ацетата, а также биосинтез белков; усиливает синтез ферментов, участвующих в глюконеогенезе; нарушает соотношение между гликолизом и глюконеогенезом.
Мышцы и печень при недостатке инсулина не способны использовать глюкозу даже при высоком ее содержании в крови. Недостаточность выделения инсулина лежит в основе возникновения заболевания— диабета. Многие последствия диабета напоминают эффект углеводного голодания, хотя содержание глюкозы в крови при этом заболевании резко повышается.
Глюкагон — гормон, продуцируемый α-клетками поджелудочной железы, активирует расщепление гликогена в печени, в результате чего освобождается глюкоза, поступающая в кровь.
Адреналин — гормон мозгового слоя надпочечников. Совместно с глюкагоном активирует гликоген-фосфорилазу печени и мышц, увеличивает содержание глюкозы в крови. Фермент встречается в двух формах — активной (фосфорилаза а) и неактивной (фосфорилаза b). Молекулярная масса фосфорилазы а составляет 380 000. Фосфорилаза а состоит из четырех субъединиц, каждая из которых содержит остаток фосфосерина и молекулу кофермента — пиридоксальфосфата, ковалентно связанную с остатком лизина. Фосфатаза фосфорилазы вызывает гидролитическое расщепление связи фосфатной группы с остатком серина и образование димеров.
Механизмы действия адреналина и глюкагона на поддержание уровня глюкозы в крови аналогичны.
Глюкокортикоиды (гормоны коркового слоя надпочечников) усиливают глюконеогенез — новообразование сахара из неуглеводов. Это приводит к увеличению количества глюкозы в крови и гликогена в печени. Соматотропный гормон гипофиза уменьшает утилизацию глюкозы периферическими тканями, усиливает глюконеогенез, угнетает продукцию инсулина, в итоге повышает содержание глюкозы в крови. Тироксин и трийодтиронин (гормоны щитовидной железы) в умеренных дозах усиливают всасывание моносахаридов в кишечнике. Избыток этих гормонов тормозит окислительное фосфорилирование и способствует повышению концентрации глюкозы в крови.
Содержание гормонов в крови находится под контролем центральной нервной системы, осуществляющей контроль за уровнем глюкозы в крови через эндокринную систему. Однако помимо опосредованного влияния на уровень глюкозы в крови центральная нервная система (ЦНС) осуществляет и прямое воздействие.
За счет контроля со стороны ЦНС и эндокринной системы в крови автоматически поддерживается постоянный уровень глюкозы, происходит саморегуляция, которая может осуществляться на различных уровнях. Так, например, кровь с избыточным содержанием глюкозы, достигая поджелудочной железы, непосредственно стимулирует выделение инсулина. Последний усиливает поглощение глюкозы тканями, использование ее на синтез гликогена, в результате концентрация глюкозы снижается до нормы.
6.8. Превращения углеводов, связанные с дыханием и брожением
Гликолиз
Стадии и реакции гликолиза. Гликолиз — это анаэробный процесс, приводящий к распаду одной молекулы глюкозы на две молекулы молочной кислоты. При этом освобождается энергия, которую организм аккумулирует в форме АТФ. Реакции гликолиза протекают в цитозоле, без потребления кислорода. В анаэробных условиях гликолиз — единственный процесс в организме животных, растений и многих бактерий, поставляющий энергию.
При аэробном расщеплении глюкозы один из конечных продуктов гликолиза — пировиноградная кислота окисляется до С02 и Н20 в цикле трикарбоновых кислот. Реакция этого цикла осуществляется в митохондриях (у бактерий — в соответствующих мембранных образованиях) при участии кислорода.
Гликолиз протекает в две стадии.
Первая стадия — подготовительная, или собирательная. На этой стадии различные гексозы вовлекаются в гликолиз. Хотя в реакциях гликолиза окисляется главным образом глюкоза, наряду с ней могут вступить в этот процесс другие различные гексозы, например фруктоза, манноза. При этом инертные молекулы гексоз активируются, фосфорилируются за счет АТФ, превращаются в глюкозо-6-фосфат. Этот этап заканчивается образованием глицеральдегид-3-фосфата.
Вторая стадия — окислительная. Глицеральдегид-З-фосфат окисляется до пировиноградной кислоты (пируват) или молочной кислоты (лактат). Энергия окисления накапливается в АТФ, образуются восстановительные эквиваленты НАДН.
Гексокиназа
АТФ + Глюкоза → АДФ + Глюкозо-6-фосфат
Эту реакцию катализируют ферменты двух типов, различающиеся по своей специфичности в отношении cахаров, — гексокиназа и глюкокиназа. Гексокиназа играет основную роль, именно она функционирует в большинстве клеток. Гексокиназа способна катализировать фосфорилирование не только D-глюкозы, но также и многих других гексоз, например D-фруктозы, D-маннозы и D-глюкозамина.
Глюкокиназа фосфорилирует только D-глюкозу, к тому же обладает к ней более низким сродством, чем гексокиназа. Глюкокиназа содержится в печени. Она вступает в действие только в том случае, когда концентрация глюкозы в крови очень высока. Обе киназы нуждаются в присутствии двухвалентных катионов (Mg24+ или Мп2+), которые связываются с АТФ, образуя истинные субстраты— MgАТФ2- или МnАТФ2-. В ходе гексокиназной реакции происходит значительное уменьшение свободной энергии, что обеспечивает необратимость этой реакции внутри клетки. Скорость реакции тормозится ее продуктом — глюкозо-6-фосфатом.
Образовавшийся при гликолизе в цитоплазме НАДН не может проникать внутрь митохондрий. В аэробных условиях он передает электроны и протон в дыхательную цепь митохондрий посредством глицерофосфатного челночного механизма. В цитоплазме НАДН реагирует с дигидроксиацетонфосфатом, образуя глицерол-3-фосфат, который легко проникает через мембрану митохондрий. Реакция катализируется цитоплазматической глицерол-3-фосфат — дегидрогеназой (НАД+):
Дигидроксиацетонфосфат + НАДН + Н+ ⇆ Глицерол-З-фосфат + НАД+
Внутри митохондрии глицерол-3-фосфат снова окисляется до дигидроксиацетонфосфата, но уже с помощью флавинового фермента— митохондриальной глицерол-3-фосфат — дегидрогеназы:
Глицерол-З-фосфат + ФАД⇆ Дигидроксиацетонфосфат + ФАДН2
Цитоплазматический НАДН восстанавливает щавелевоуксусную кислоту до яблочной, легко проникающей в митохондрии и там снова окисляющейся до щавелевоуксусной кислоты с передачей водорода в дыхательную цепь.
У растений в некоторых тканях гликолиз протекает даже при достаточном доступе кислорода. Особенно характерно такое однофазовое, «анаэробное дыхание» (или «аэробное брожение») для зародышевой ткани семян и внутренних тканей плодов растений вследствие недостатка в них кислорода. Гликолиз является основным путем катаболизма глюкозы гомоферментативных молочнокислых бактерий, дрожжей.
Брожение. Многие микроорганизмы (дрожжевые грибы, плесневые грибы, бактерии) получают энергию за счет брожения. Это наиболее примитивный способ получения энергии. Разнообразные брожения совпадают с гликолизом во многих своих стадиях.
Часто встречающийся тип брожения — молочнокислое. В результате гомоферментативного молочнокислого брожения из глюкозы образуется молочная кислота с почти 100%-ным выходом. Реакции этого процесса идентичны реакциям гликолиза. При гетероферментативном (смешанное) молочнокислом брожении из глюкозы кроме молочной кислоты образуются в значительном количестве другие продукты — уксусная кислота, этиловый спирт и оксид углерода (IV). Химизм этого процесса совершенно иной, в основе его лежит окисление глюкозы по пентозофосфатному пути.
Молочнокислое брожение протекает при силосовании кормов для сельскохозяйственных животных, квашении капусты. Образующаяся молочная кислота предотвращает развитие гнилостных бактерий, плесневых грибов, т. е. служит консервантом.
Пропионовая кислота представляет собой продукт сбраживания углеводов пропионовыми бактериями. Наряду с ней образуются уксусная кислота и оксид углерода (IV). На каждую молекулу ПВК, окисленную до уксусной кислоты и С02, приходится две молекулы ПВК, которые превращаются в пропионовую кислоту.
Уксуснокислые бактерии (рода Acetobacter) окисляют этиловый спирт до уксусной кислоты. Эти бактерии способны окислять и другие спирты, в том числе предельные многоатомные. Такое явление называется неполным окислением.
6.9. Глюконеогенез.
Анаэробная фаза расщепления глюкозы — гликолиз — заканчивается образованием ПВК или молочной кислоты. Они при определенных условиях могут вновь ресинтезироваться в глюкозу. Из двух молекул молочной кислоты образуется одна молекула глюкозы, т. е. происходит как бы обращение гликолиза. Этот процесс называется глюконеогенезом. Если гликолиз — центральный путь катаболизма углеводов, то глюконеогенез — анаболический процесс, наиболее важный общий путь биосинтеза моносахаридов и полисахаридов у человека, животных, многих бактерий. У фотосинтезирующих организмов он играет обычно второстепенную роль.
Большинство стадий глюконеогенеза представляет собой обращение реакций гликолиза. Однако существуют три необратимые стадии гликолиза, протекающие с выделением значительного количества энергии, поэтому глюконеогенез идет в обход этих стадий.
Добавить комментарий